

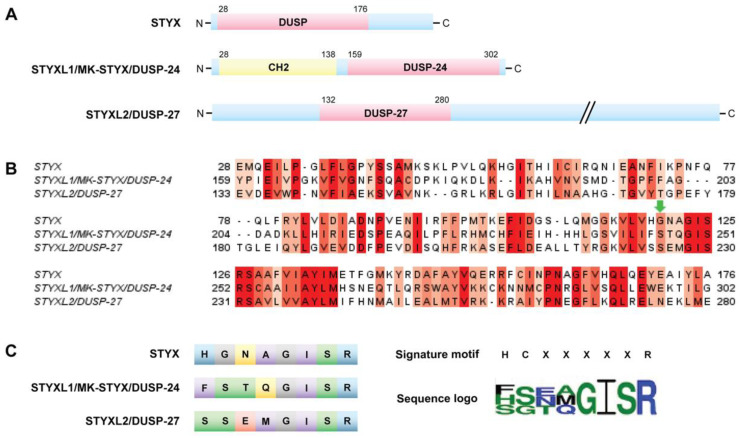
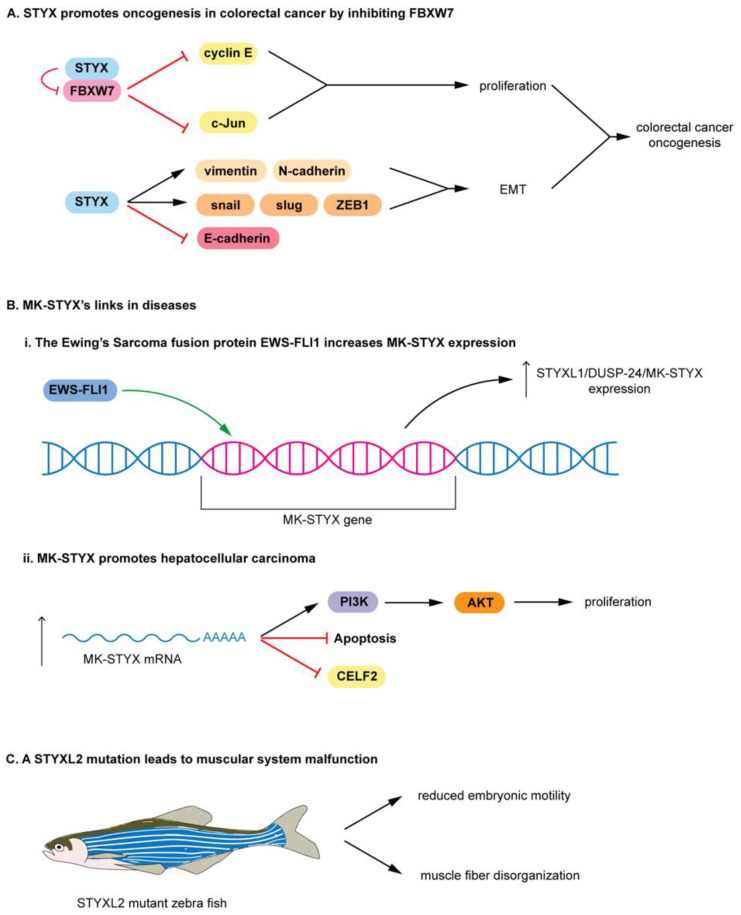
The Roles of Pseudophosphatases in Disease
- PMID: 34203203
- PMCID: PMC8269279
- DOI: 10.3390/ijms22136924
The Roles of Pseudophosphatases in Disease
Abstract
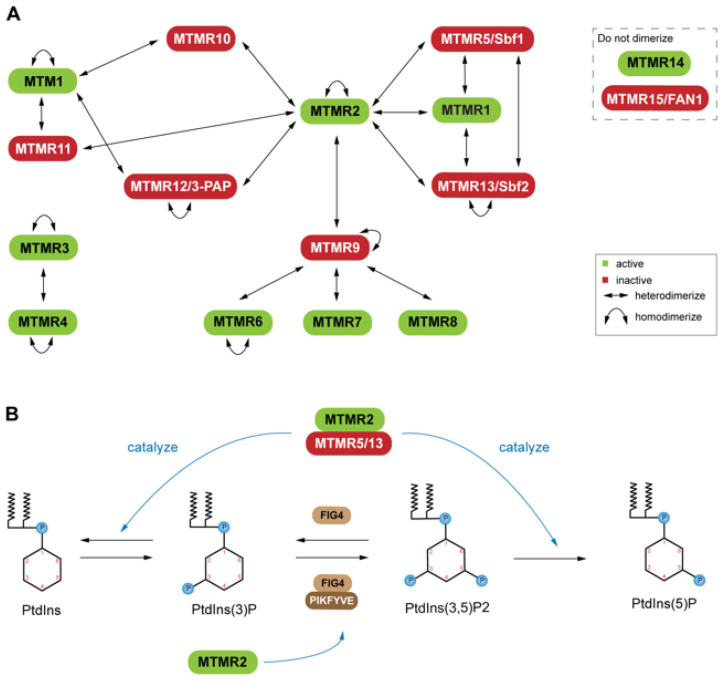
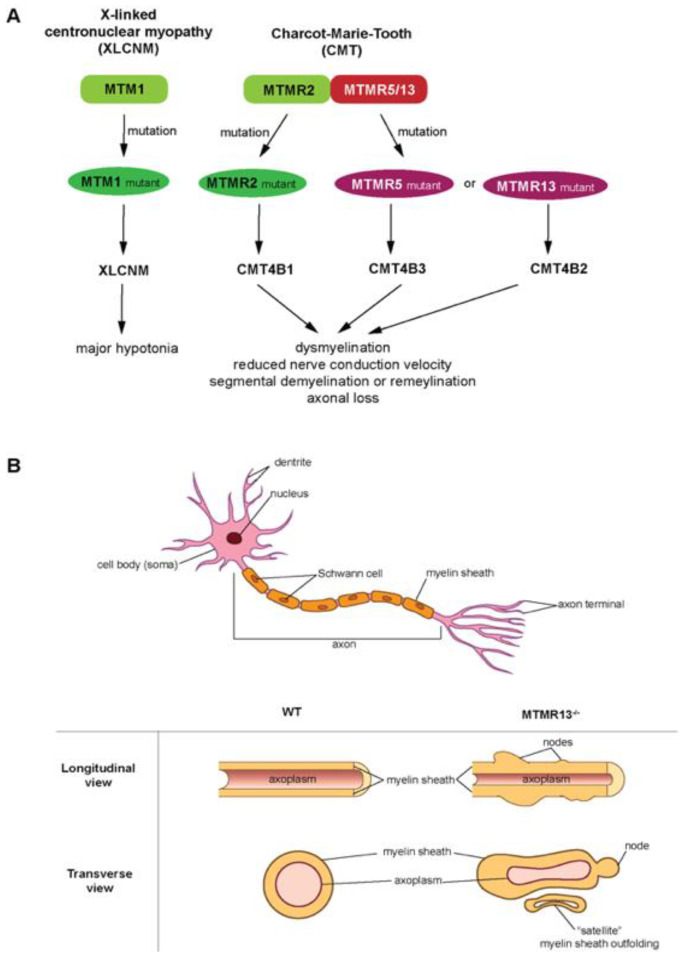
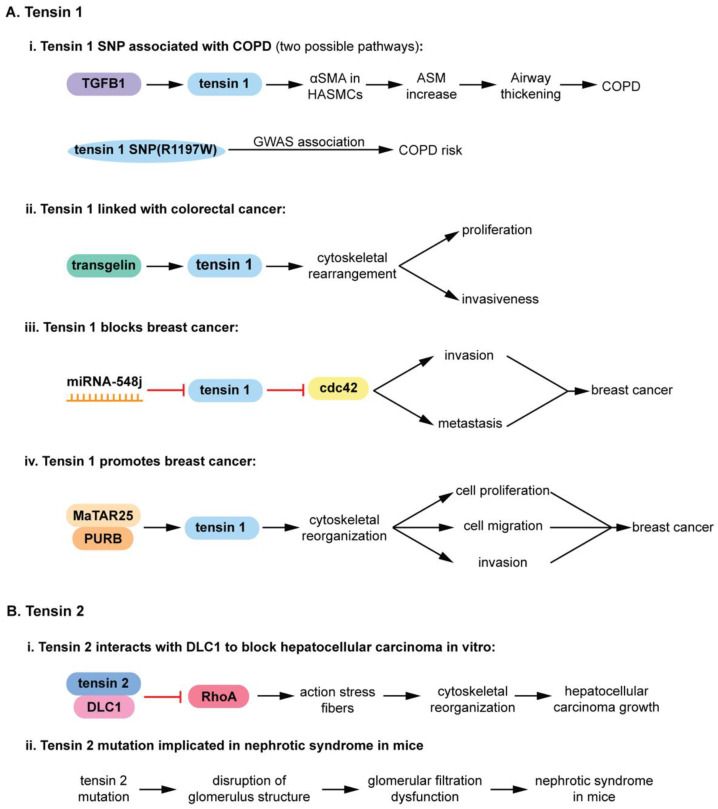
The pseudophosphatases, atypical members of the protein tyrosine phosphatase family, have emerged as bona fide signaling regulators within the past two decades. Their roles as regulators have led to a renaissance of the pseudophosphatase and pseudoenyme fields, catapulting interest from a mere curiosity to intriguing and relevant proteins to investigate. Pseudophosphatases make up approximately fourteen percent of the phosphatase family, and are conserved throughout evolution. Pseudophosphatases, along with pseudokinases, are important players in physiology and pathophysiology. These atypical members of the protein tyrosine phosphatase and protein tyrosine kinase superfamily, respectively, are rendered catalytically inactive through mutations within their catalytic active signature motif and/or other important domains required for catalysis. This new interest in the pursuit of the relevant functions of these proteins has resulted in an elucidation of their roles in signaling cascades and diseases. There is a rapid accumulation of knowledge of diseases linked to their dysregulation, such as neuropathies and various cancers. This review analyzes the involvement of pseudophosphatases in diseases, highlighting the function of various role(s) of pseudophosphatases involvement in pathologies, and thus providing a platform to strongly consider them as key therapeutic drug targets.
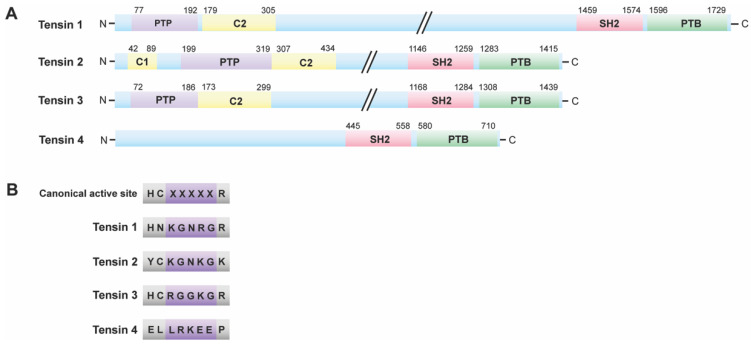
Keywords: MK-STYX (MAPK (mitogen-activated protein kinase) phosphoserine/threonine/tyrosine-binding protein); STYX (phosphoserine/threonine/tyrosine-interacting protein); disease; dual specificity phosphatases (DUSPs); myotubularin phosphatases (MTMs); protein tyrosine phosphatases (PTPs); pseudoenzymes; pseudophosphatases; tensin.
Conflict of interest statement
The authors declare no conflict of interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
