

The Crazy Ovary
- PMID: 34207147
- PMCID: PMC8234655
- DOI: 10.3390/genes12060928
The Crazy Ovary
Abstract
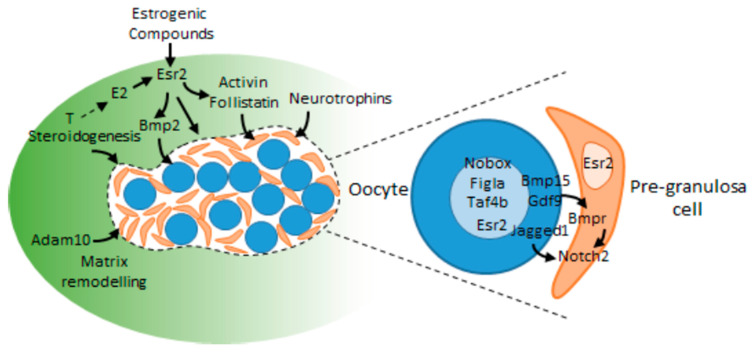
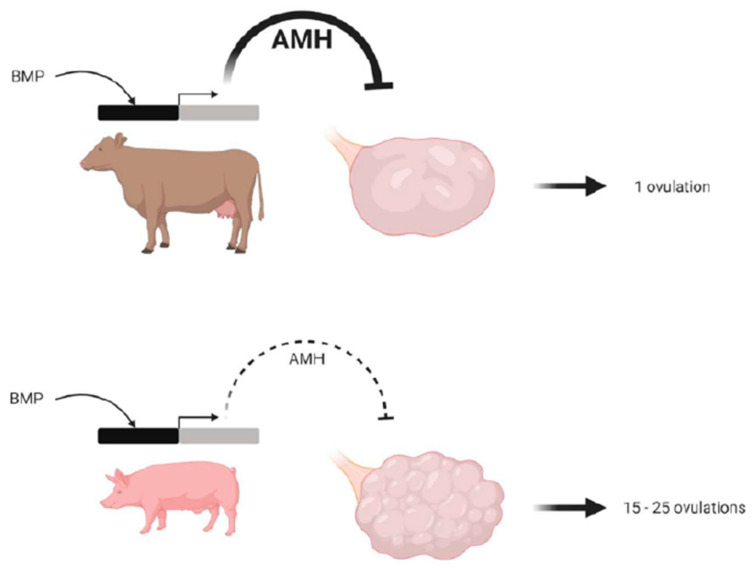
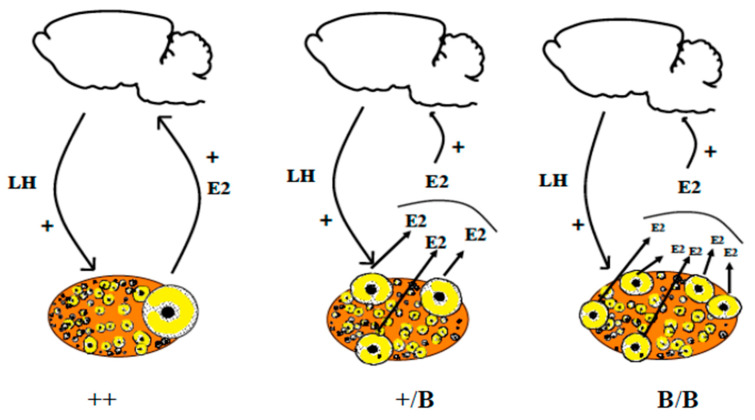
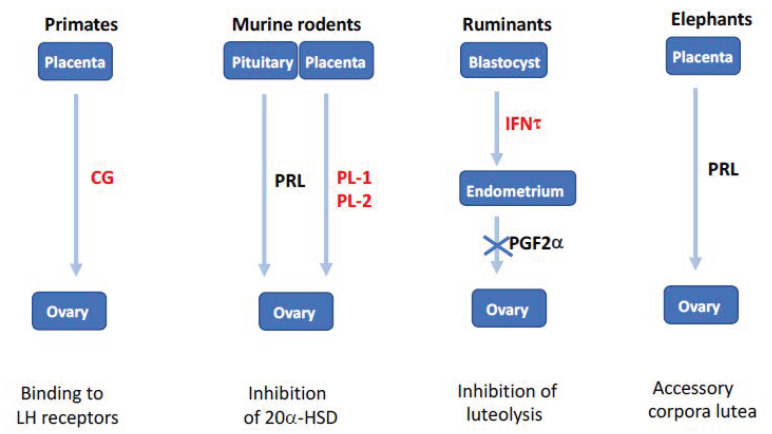
From fetal life until senescence, the ovary is an extremely active tissue undergoing continuous structural and functional changes. These ever-changing events are best summarized by a quotation attributed to Plato when describing motion in space and time-'nothing ever is but is always becoming…'. With respect to the ovary, these changes include, at the beginning, the processes of follicular formation and thereafter those of follicular growth and atresia, steroidogenesis, oocyte maturation, and decisions relating to the number of mature oocytes that are ovulated for fertilization and the role of the corpus luteum. The aims of this review are to offer some examples of these complex and hitherto unknown processes. The ones herein have been elucidated from studies undertaken in vitro or from normal in vivo events, natural genetic mutations or after experimental inactivation of gene function. Specifically, this review offers insights concerning the initiation of follicular growth, pathologies relating to poly-ovular follicles, the consequences of premature loss of germ cells or oocytes loss, the roles of AMH (anti-Müllerian hormone) and BMP (bone morphogenetic protein) genes in regulating follicular growth and ovulation rate together with species differences in maintaining luteal function during pregnancy. Collectively, the evidence suggests that the oocyte is a key organizer of normal ovarian function. It has been shown to influence the phenotype of the adjacent somatic cells, the growth and maturation of the follicle, and to determine the ovulation rate. When germ cells or oocytes are lost prematurely, the ovary becomes disorganized and a wide range of pathologies may arise.
Keywords: crazy; folliculogenesis; ovary; ovulation rate; primordial follicles; reactivation; steroidogenesis; unexpected.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
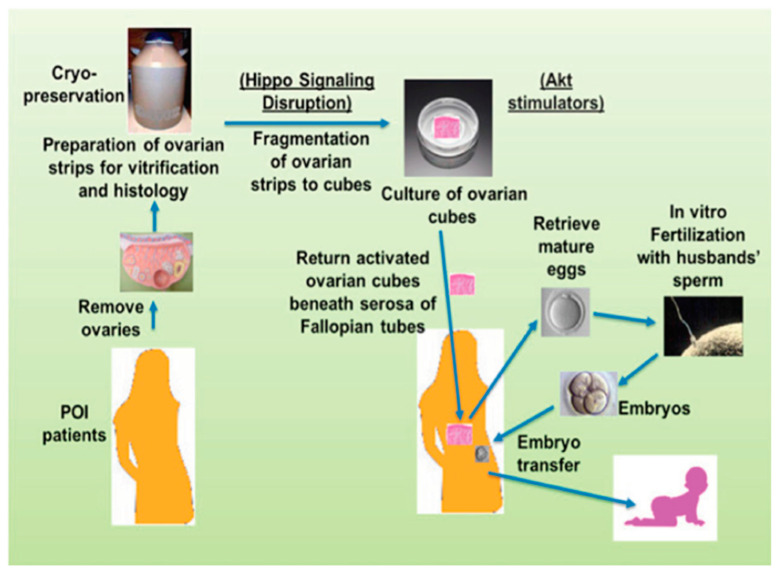
- Kawamura K., Cheng Y., Suzuki N., Deguchi M., Sato Y., Takae S., Ho C.H., Kawamura N., Tamura M., Hashimoto S., et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA. 2013;110:17474–17479. doi: 10.1073/pnas.1312830110. - DOI - PMC - PubMed
-
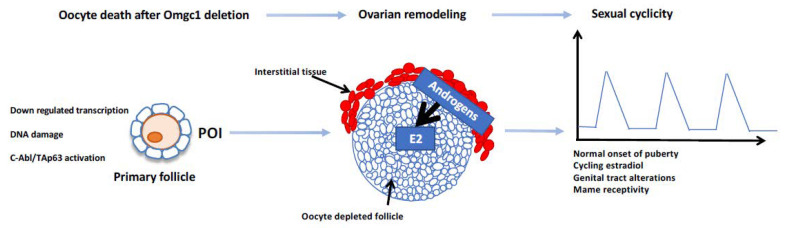
- Vandormael-Pournin S., Guigon C.J., Ishaq M., Coudouel N., Ave P., Huerre M., Magre S., Cohen-Tannoudji J., Cohen-Tannoudji M. Oocyte-specific inactivation of Omcg1 leads to DNA damage and c-Abl/TAp63-dependent oocyte death associated with dramatic remodeling of ovarian somatic cells. Cell Death Differ. 2015;22:108–117. doi: 10.1038/cdd.2014.122. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
