

Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin
- PMID: 34209313
- PMCID: PMC8307532
- DOI: 10.3390/md19070380
Electrophysiological Evaluation of Pacific Oyster (Crassostrea gigas) Sensitivity to Saxitoxin and Tetrodotoxin
Abstract
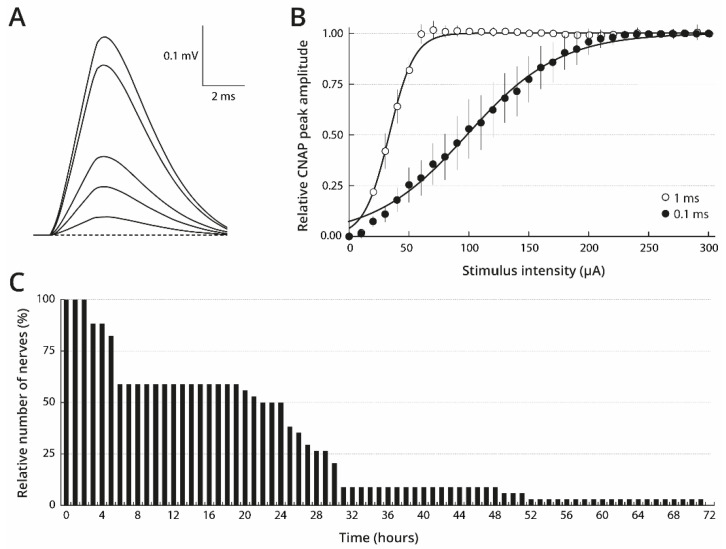
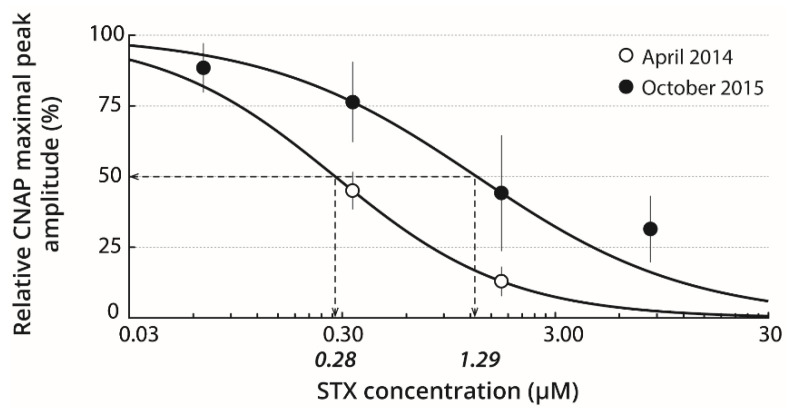
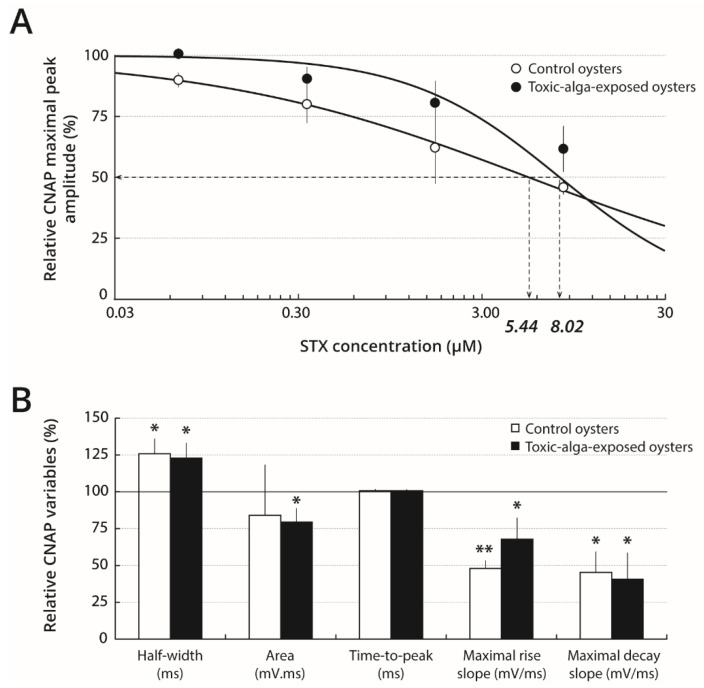
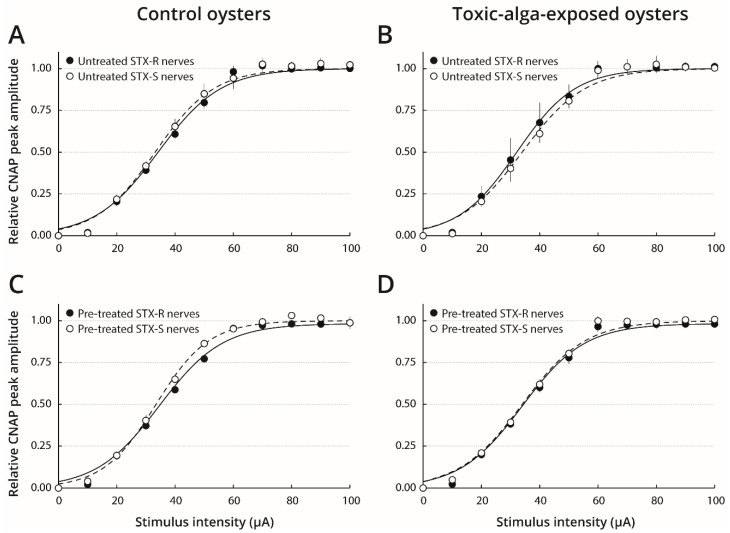
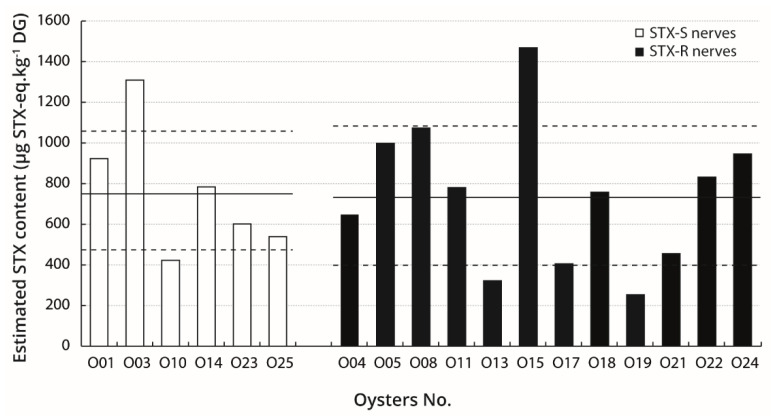
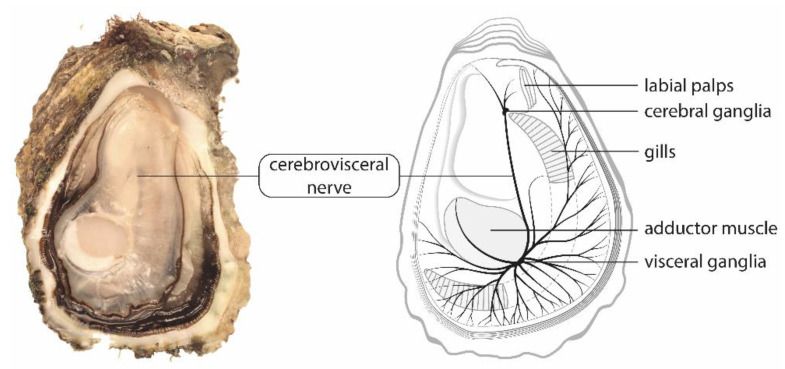
Pacific oysters (Crassostrea gigas) may bio-accumulate high levels of paralytic shellfish toxins (PST) during harmful algal blooms of the genus Alexandrium. These blooms regularly occur in coastal waters, affecting oyster health and marketability. The aim of our study was to analyse the PST-sensitivity of nerves of Pacific oysters in relation with toxin bio-accumulation. The results show that C. gigas nerves have micromolar range of saxitoxin (STX) sensitivity, thus providing intermediate STX sensitivity compared to other bivalve species. However, theses nerves were much less sensitive to tetrodotoxin. The STX-sensitivity of compound nerve action potential (CNAP) recorded from oysters experimentally fed with Alexandrium minutum (toxic-alga-exposed oysters), or Tisochrysis lutea, a non-toxic microalga (control oysters), revealed that oysters could be separated into STX-resistant and STX-sensitive categories, regardless of the diet. Moreover, the percentage of toxin-sensitive nerves was lower, and the STX concentration necessary to inhibit 50% of CNAP higher, in recently toxic-alga-exposed oysters than in control bivalves. However, no obvious correlation was observed between nerve sensitivity to STX and the STX content in oyster digestive glands. None of the nerves isolated from wild and farmed oysters was detected to be sensitive to tetrodotoxin. In conclusion, this study highlights the good potential of cerebrovisceral nerves of Pacific oysters for electrophysiological and pharmacological studies. In addition, this study shows, for the first time, that C. gigas nerves have micromolar range of STX sensitivity. The STX sensitivity decreases, at least temporary, upon recent oyster exposure to dinoflagellates producing PST under natural, but not experimental environment.
Keywords: Alexandrium minutum; Crassostrea gigas; compound nerve action potential; paralytic shellfish toxins.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Lassus P., Baron R., Garen P., Truquet P., Masselin P., Bardouil M., Leguay D., Amzil Z. Paralytic shellfish poison outbreaks in the Penzé estuary: Environmental factors affecting toxin uptake in the oyster. Crassostrea gigas. Aquat. Living Resour. 2004;17:207–214. doi: 10.1051/alr:2004012. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
