

Spindle assembly checkpoint activation and silencing at kinetochores
- PMID: 34210579
- PMCID: PMC8406419
- DOI: 10.1016/j.semcdb.2021.06.009
Spindle assembly checkpoint activation and silencing at kinetochores
Abstract
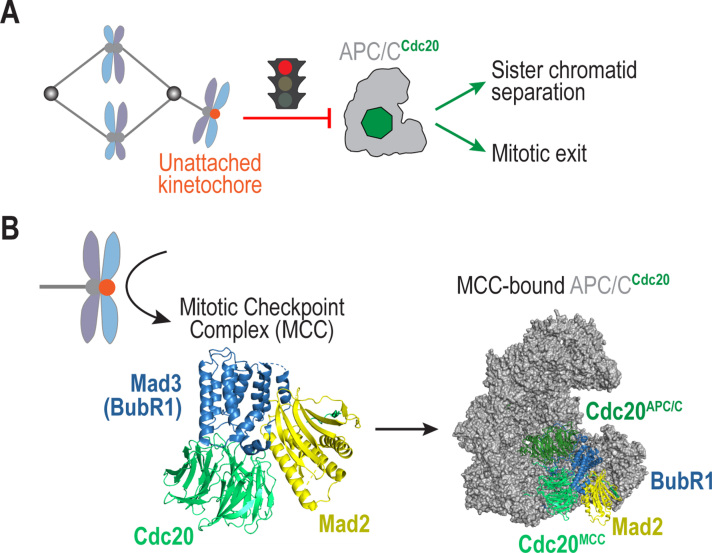
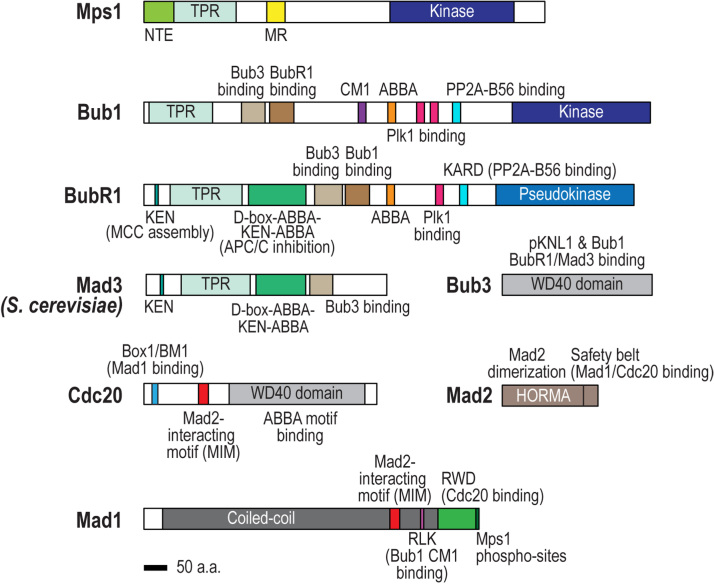
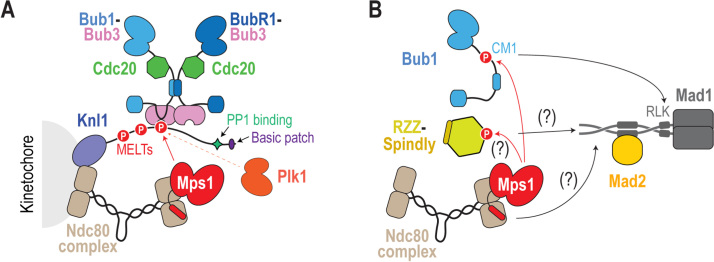
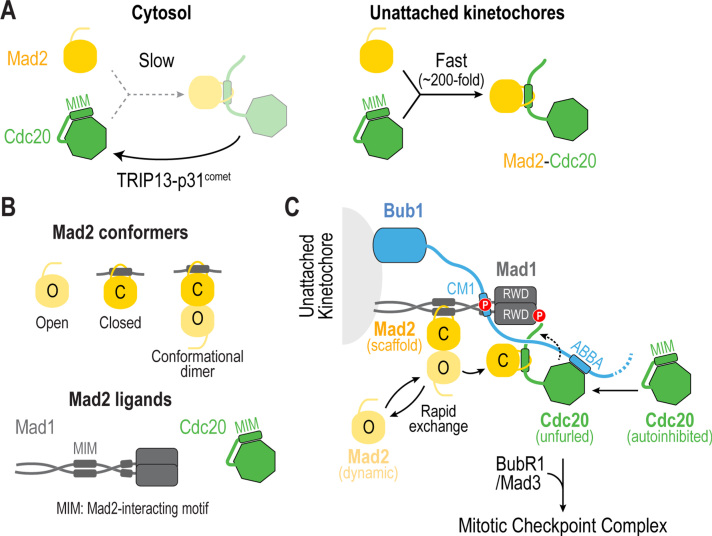
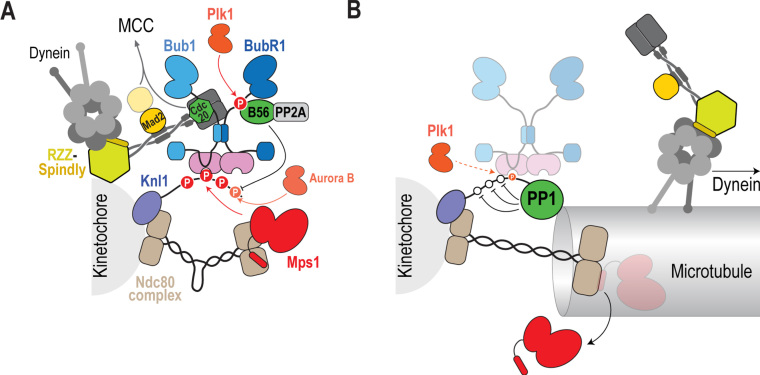
The spindle assembly checkpoint (SAC) is a surveillance mechanism that promotes accurate chromosome segregation in mitosis. The checkpoint senses the attachment state of kinetochores, the proteinaceous structures that assemble onto chromosomes in mitosis in order to mediate their interaction with spindle microtubules. When unattached, kinetochores generate a diffusible inhibitor that blocks the activity of the anaphase-promoting complex/cyclosome (APC/C), an E3 ubiquitin ligase required for sister chromatid separation and exit from mitosis. Work from the past decade has greatly illuminated our understanding of the mechanisms by which the diffusible inhibitor is assembled and how it inhibits the APC/C. However, less is understood about how SAC proteins are recruited to kinetochores in the absence of microtubule attachment, how the kinetochore catalyzes formation of the diffusible inhibitor, and how attachments silence the SAC at the kinetochore. Here, we summarize current understanding of the mechanisms that activate and silence the SAC at kinetochores and highlight open questions for future investigation.
Keywords: Aneuploidy; Catalysis; Checkpoint; Chromosome segregation; Kinetochores; Mitosis.
Copyright © 2021. Published by Elsevier Ltd.
Figures
References
-
- Lara-Gonzalez P., Westhorpe F.G., Taylor S.S. The spindle assembly checkpoint. Curr. Biol. 2012;22(22):R966–R980. - PubMed
-
- Jia L., Kim S., Yu H. Tracking spindle checkpoint signals from kinetochores to APC/C. Trends Biochem. Sci. 2013;38(6):302–311. - PubMed
-
- Musacchio A. The molecular biology of spindle assembly checkpoint signaling dynamics. Curr. Biol. 2015;25(20):R1002–R1018. - PubMed
-
- Corbett K.D. Molecular mechanisms of spindle assembly checkpoint activation and silencing. Prog. Mol. Subcell. Biol. 2017;56:429–455. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
