

Transcriptomic Analysis and C-Terminal Epitope Tagging Reveal Differential Processing and Signaling of Endogenous TLR3 and TLR7
- PMID: 34211474
- PMCID: PMC8240634
- DOI: 10.3389/fimmu.2021.686060
Transcriptomic Analysis and C-Terminal Epitope Tagging Reveal Differential Processing and Signaling of Endogenous TLR3 and TLR7
Abstract
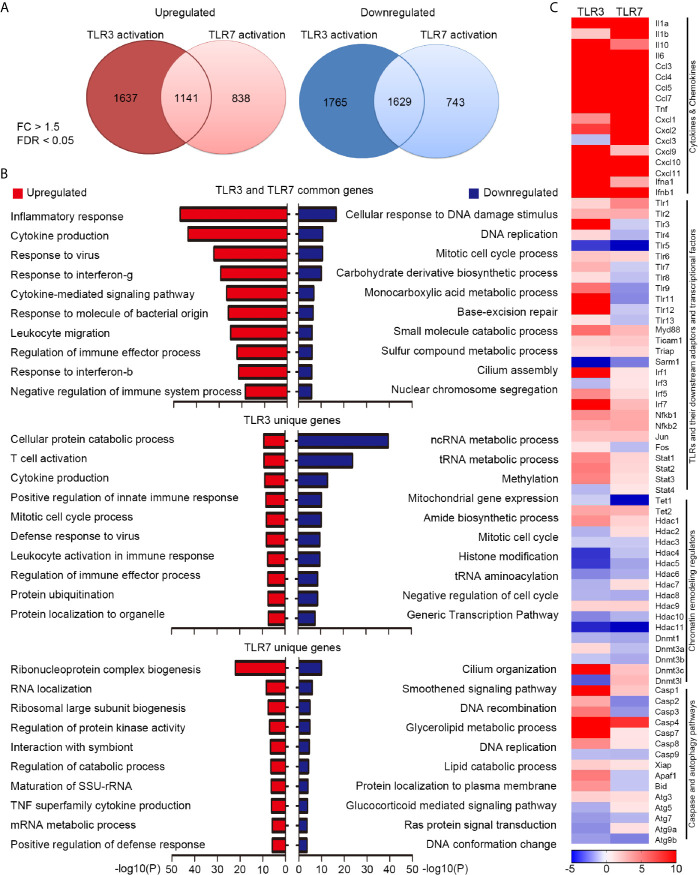
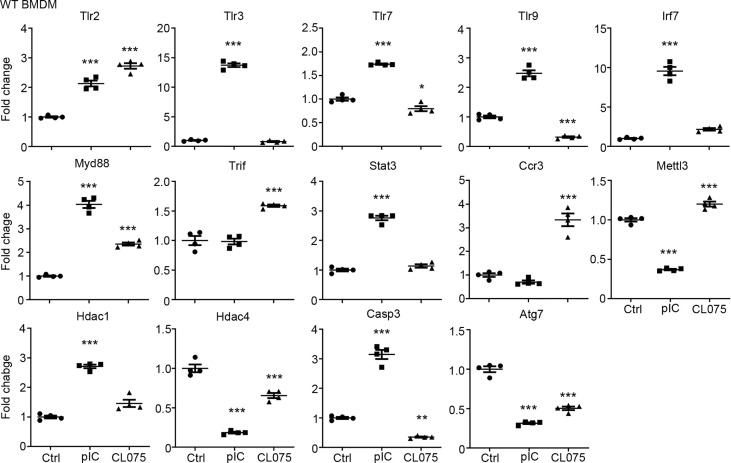
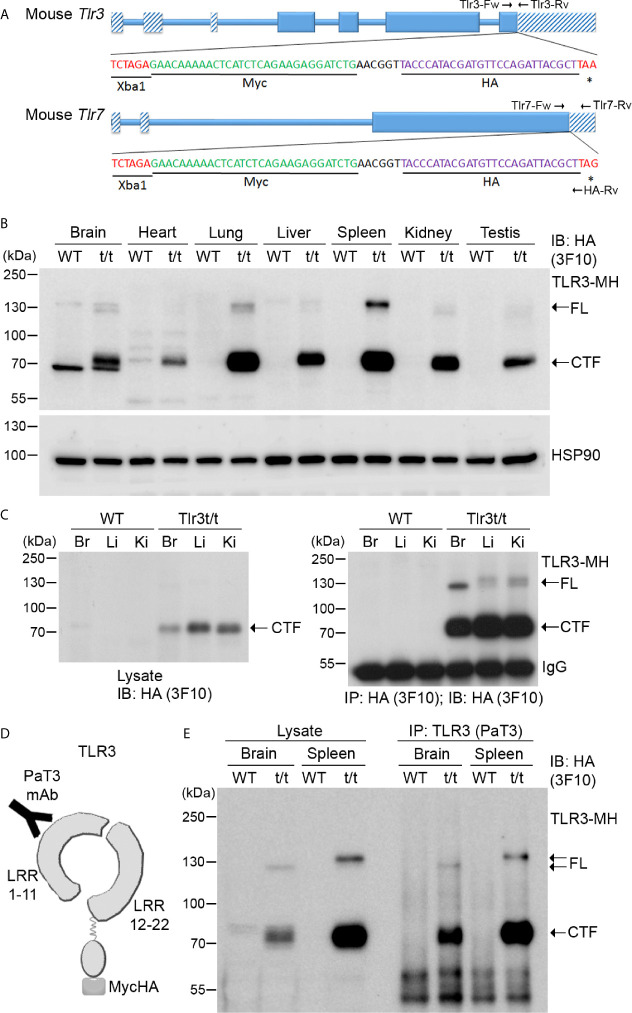
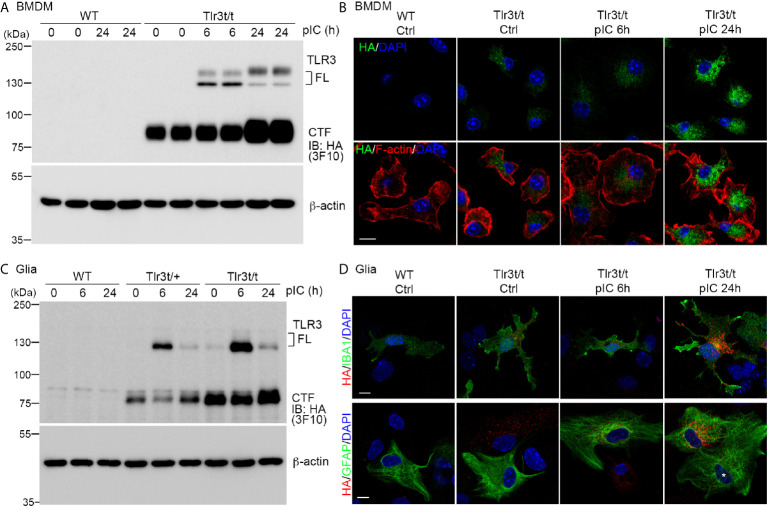
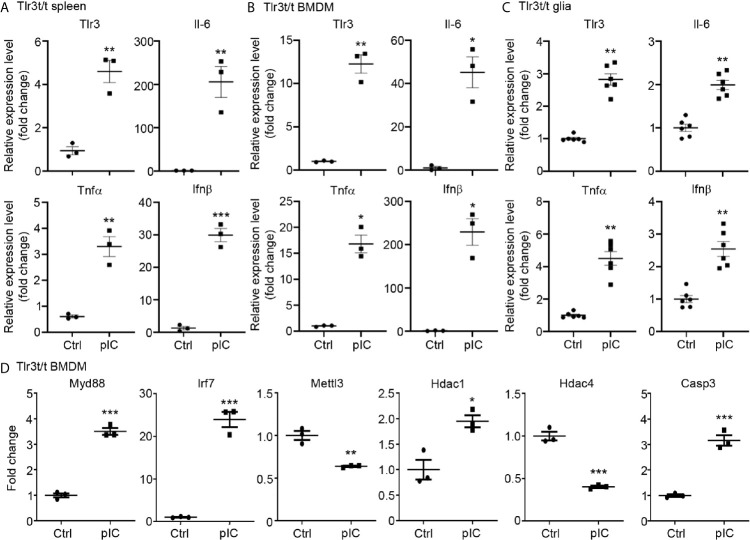
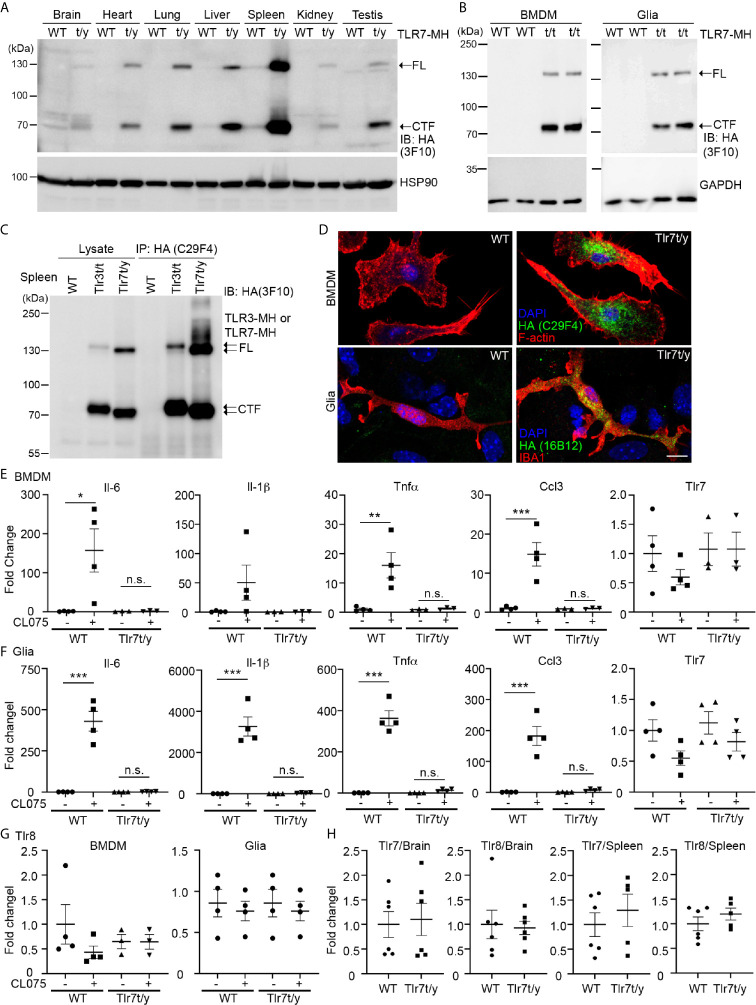
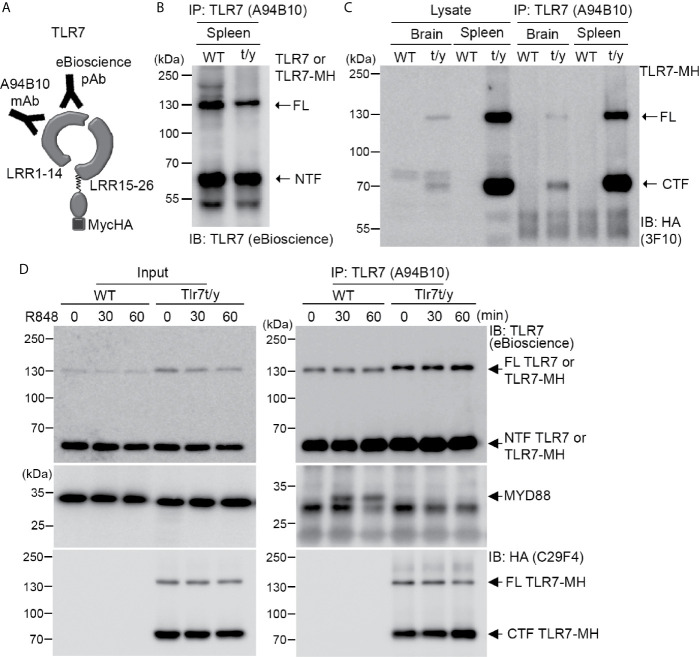
Toll-like receptor (TLR) signaling is critical for defense against pathogenic infection, as well as for modulating tissue development. Activation of different TLRs triggers common inflammatory responses such as cytokine induction. Here, we reveal differential impacts of TLR3 and TLR7 signaling on transcriptomic profiles in bone marrow-derived macrophages (BMDMs). Apart from self-regulation, TLR3, but not TLR7, induced expression of other TLRs, suggesting that TLR3 activation globally enhances innate immunity. Moreover, we observed diverse influences of TLR3 and TLR7 signaling on genes involved in methylation, caspase and autophagy pathways. We compared endogenous TLR3 and TLR7 by using CRISPR/Cas9 technology to knock in a dual Myc-HA tag at the 3' ends of mouse Tlr3 and Tlr7. Using anti-HA antibodies to detect endogenous tagged TLR3 and TLR7, we found that both TLRs display differential tissue expression and posttranslational modifications. C-terminal tagging did not impair TLR3 activity. However, it disrupted the interaction between TLR7 and myeloid differentiation primary response 88 (MYD88), the Tir domain-containing adaptor of TLR7, which blocked its downstream signaling necessary to trigger cytokine and chemokine expression. Our study demonstrates different properties for TLR3 and TLR7, and also provides useful mouse models for further investigation of these two RNA-sensing TLRs.
Keywords: MYD88; RNA-seq; epitope tagging; signalosome; toll-like receptor; transcriptomic analysis.
Copyright © 2021 Chen, Hung, Tsai, Shih, Chou, Lai, Wang and Hsueh.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials