

Commensal microbiota regulates skin barrier function and repair via signaling through the aryl hydrocarbon receptor
- PMID: 34214492
- PMCID: PMC8364505
- DOI: 10.1016/j.chom.2021.05.011
Commensal microbiota regulates skin barrier function and repair via signaling through the aryl hydrocarbon receptor
Abstract
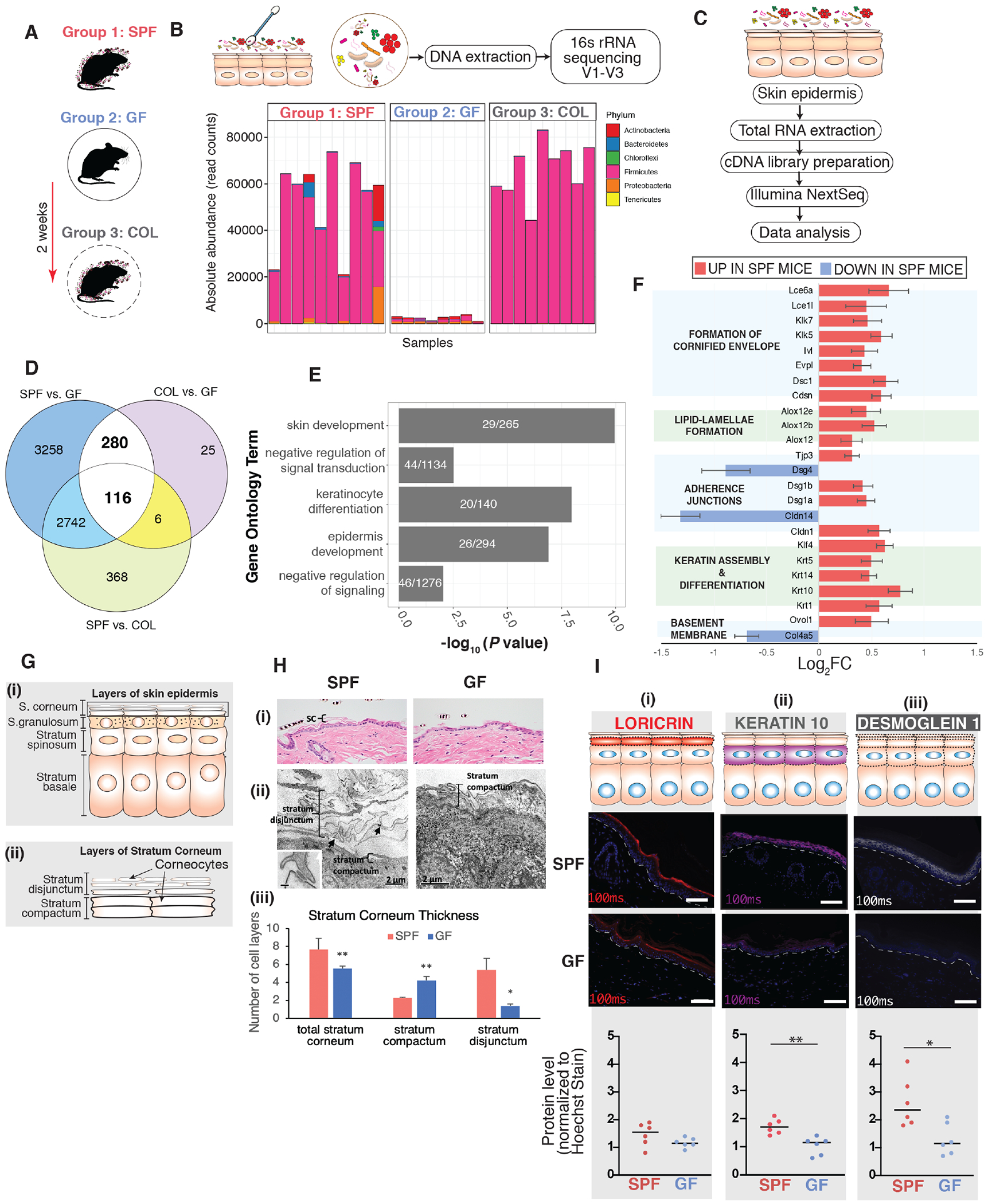
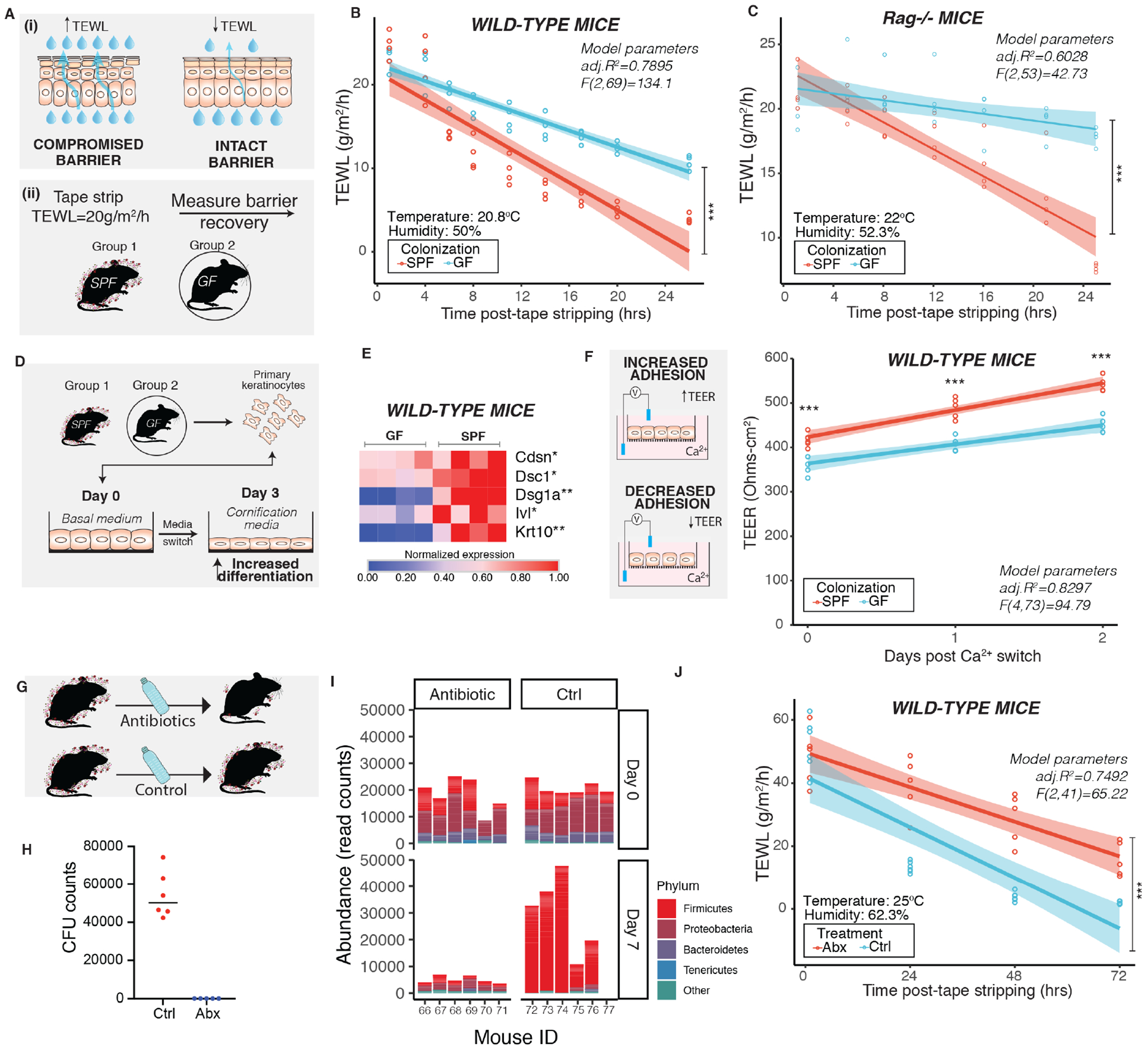
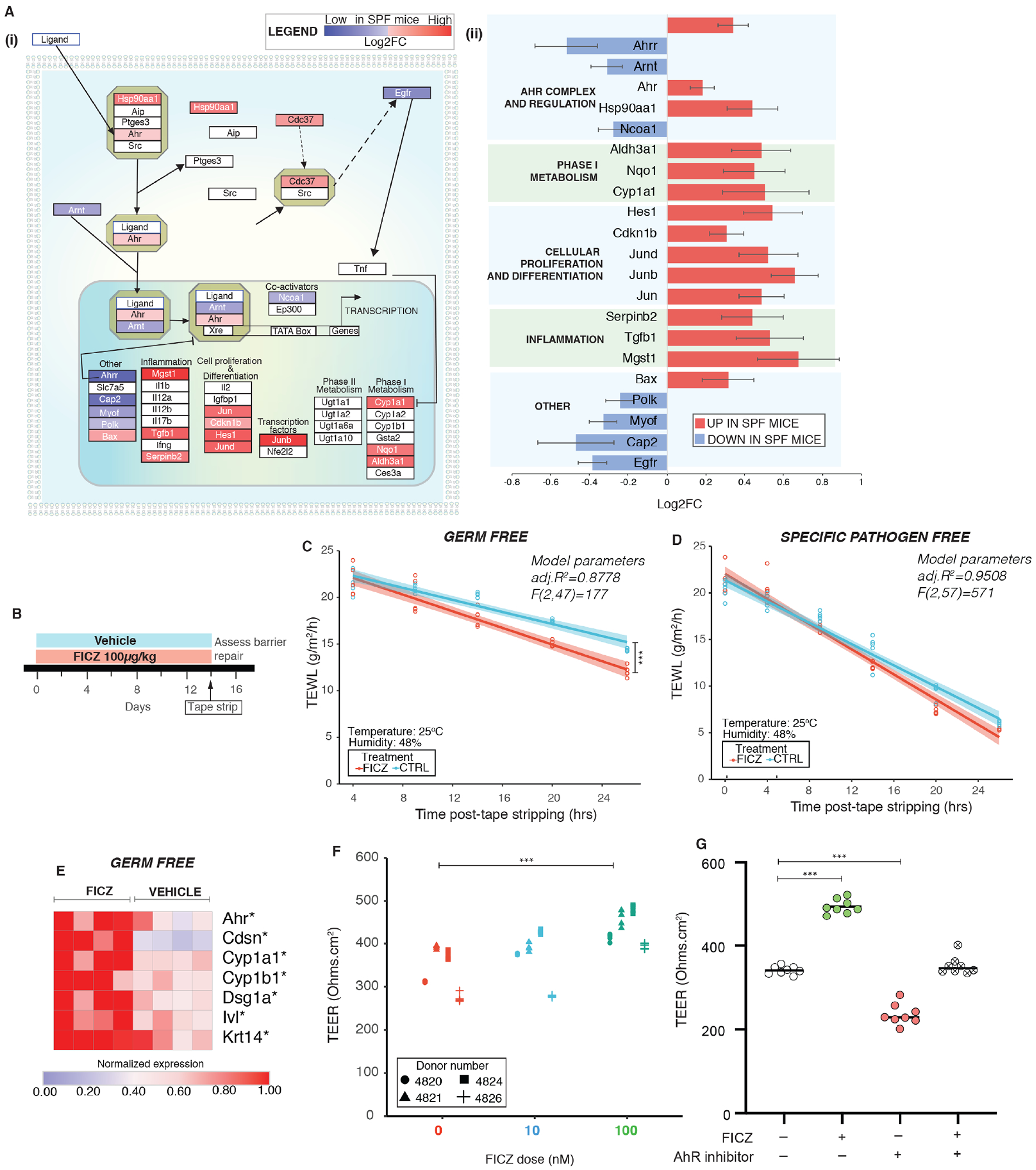
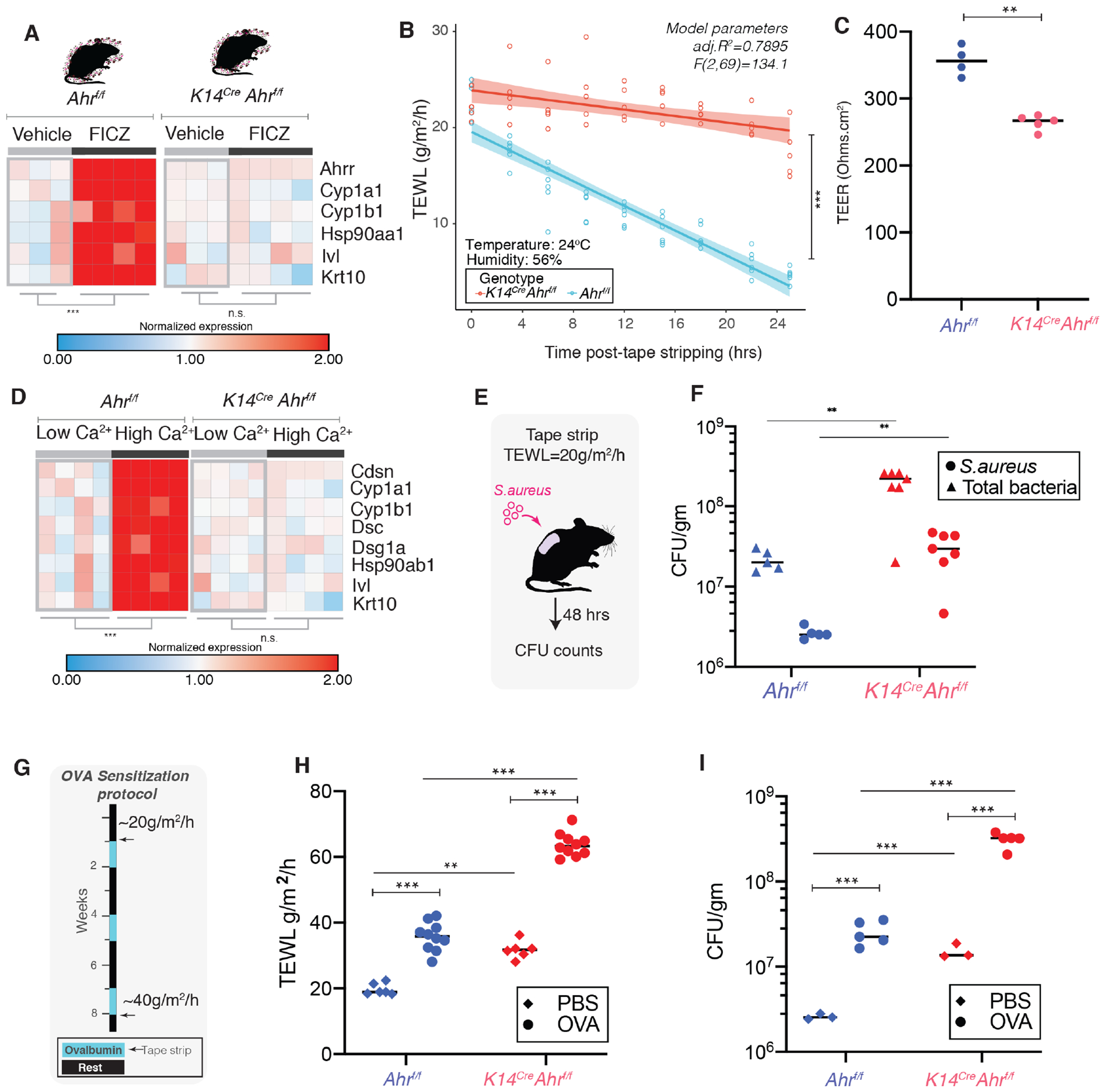
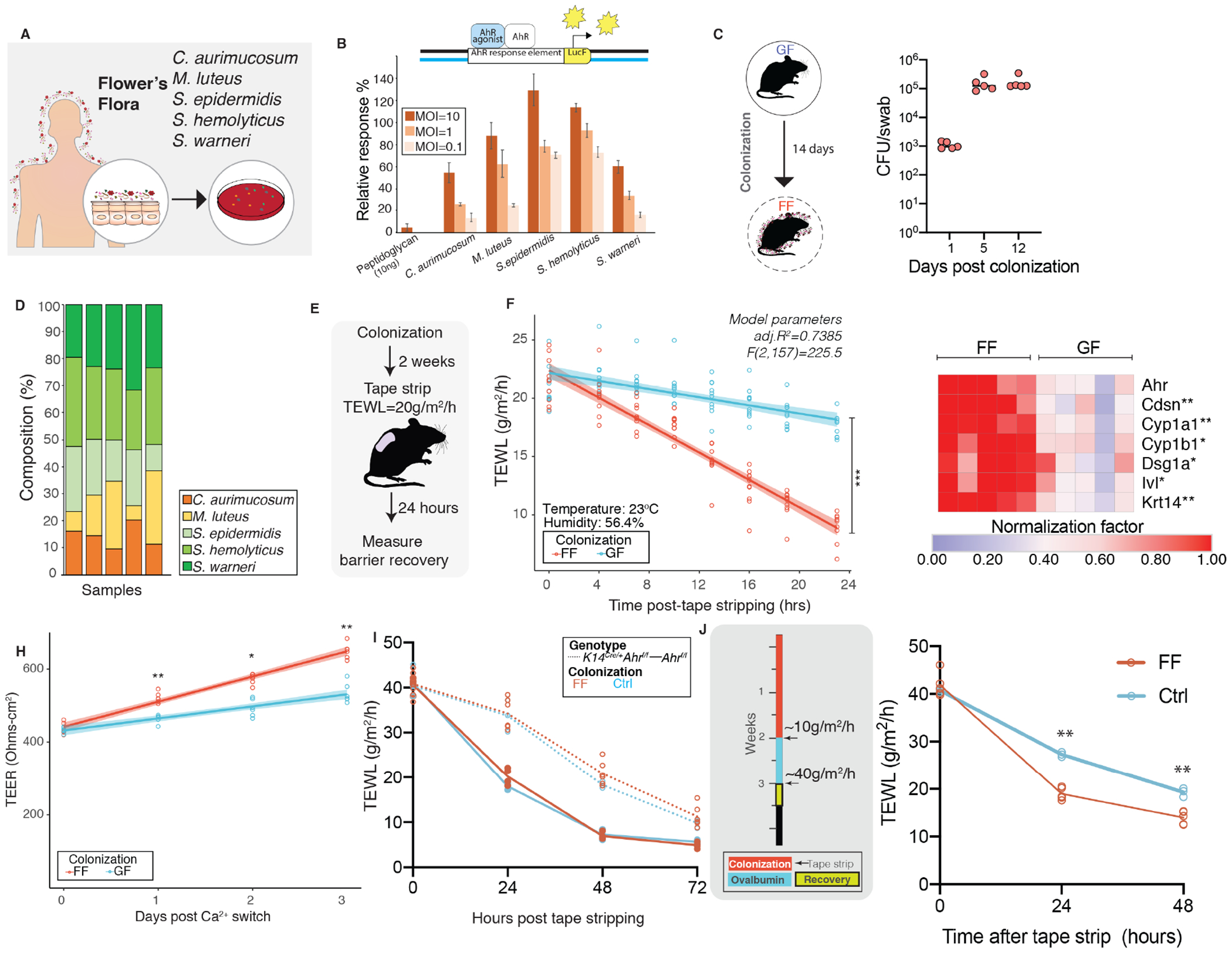
The epidermis forms a barrier that defends the body from desiccation and entry of harmful substances, while also sensing and integrating environmental signals. The tightly orchestrated cellular changes needed for the formation and maintenance of this epidermal barrier occur in the context of the skin microbiome. Using germ-free mice, we demonstrate the microbiota is necessary for proper differentiation and repair of the epidermal barrier. These effects are mediated by microbiota signaling through the aryl hydrocarbon receptor (AHR) in keratinocytes, a xenobiotic receptor also implicated in epidermal differentiation. Mice lacking keratinocyte AHR are more susceptible to barrier damage and infection, during steady-state and epicutaneous sensitization. Colonization with a defined consortium of human skin isolates restored barrier competence in an AHR-dependent manner. We reveal a fundamental mechanism whereby the microbiota regulates skin barrier formation and repair, which has far-reaching implications for the numerous skin disorders characterized by epidermal barrier dysfunction.
Keywords: aryl hydrocarbon receptor; epidermis; keratinocyte; microbiota; skin barrier; skin commensals; skin microbiome.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors have no competing interests to declare.
Figures
Comment in
-
Guardians of the barrier: Microbiota engage AHR in keratinocytes to mantain skin homeostasis.Cell Host Microbe. 2021 Aug 11;29(8):1213-1216. doi: 10.1016/j.chom.2021.07.007. Cell Host Microbe. 2021. PMID: 34384523
References
-
- Alexander H, Paller AS, Traidl-Hoffmann C, Beck LA, De Benedetto A, Dhar S, Girolomoni G, Irvine AD, Spuls P, Su J, et al. (2020). The role of bacterial skin infections in atopic dermatitis: expert statement and review from the International Eczema Council Skin Infection Group. Br J Dermatol 182, 1331–1342. - PMC - PubMed
-
- Alexa A RJ, (2020). topGO: Enrichment Analysis for Gene Ontology. In R package version 2420.
-
- Andreola F, Fernandez-Salguero PM, Chiantore MV, Petkovich MP, Gonzalez FJ, and De Luca LM (1997). Aryl hydrocarbon receptor knockout mice (AHR−/−) exhibit liver retinoid accumulation and reduced retinoic acid metabolism. Cancer Res 57, 2835–2838. - PubMed
-
- Belkaid Y, and Segre JA (2014). Dialogue between skin microbiota and immunity. Science 346, 954–959. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
