

Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance
- PMID: 34215692
- PMCID: PMC8271637
- DOI: 10.1073/pnas.2026152118
Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance
Abstract
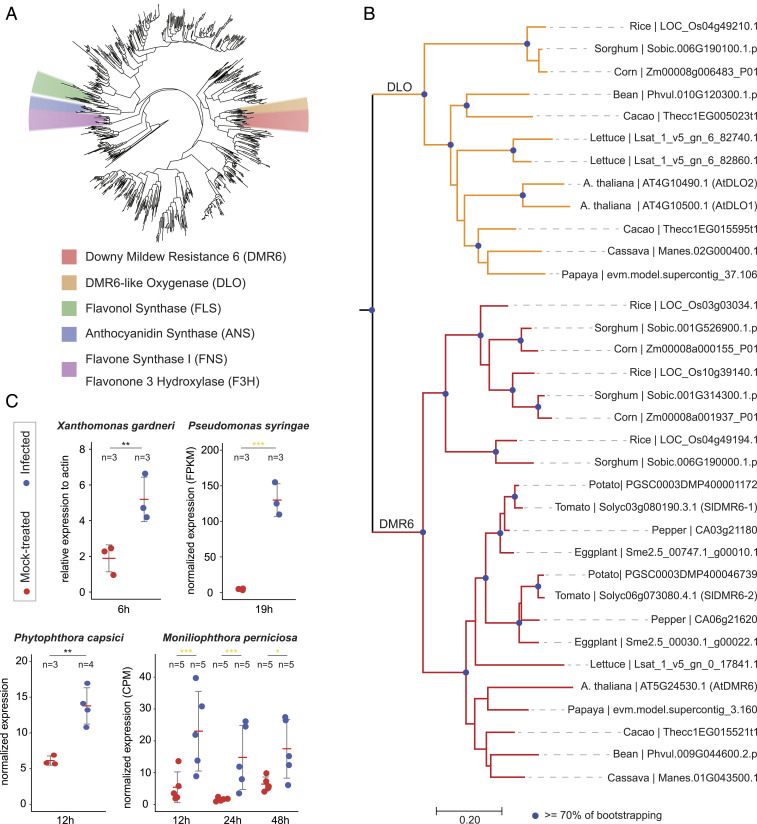
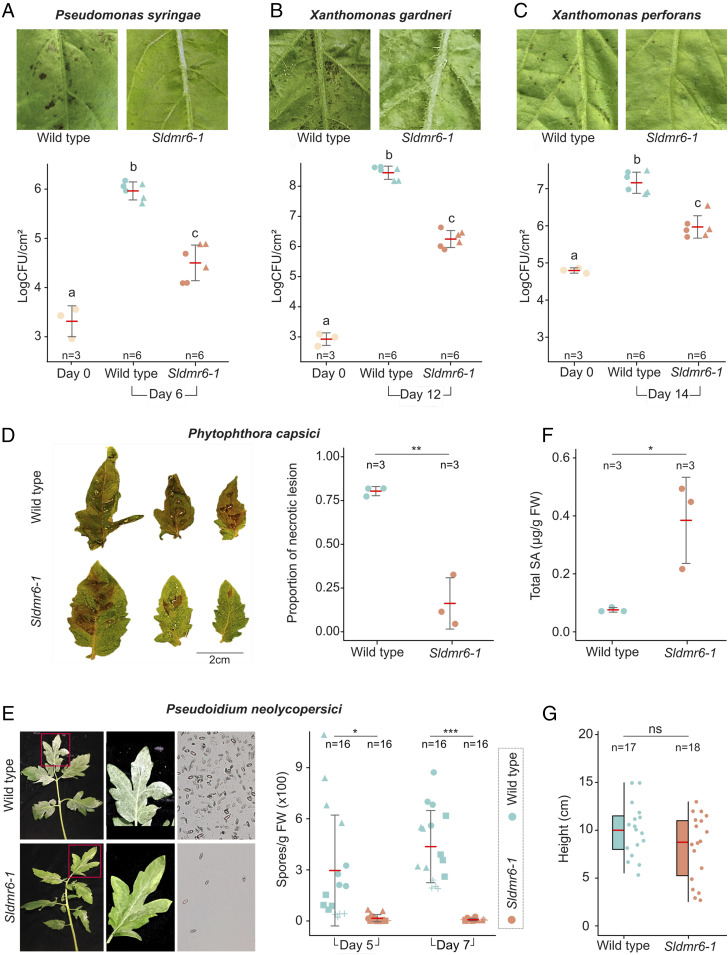
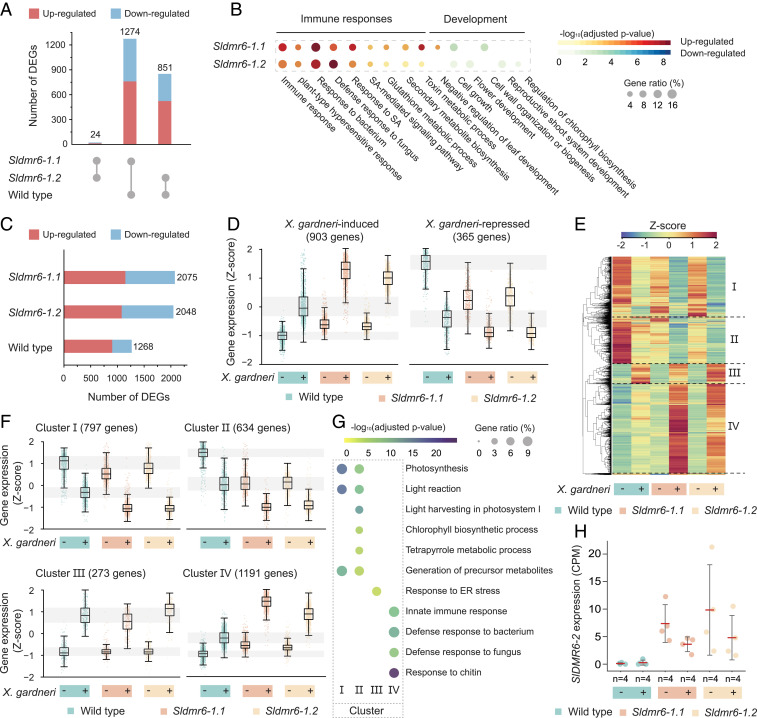
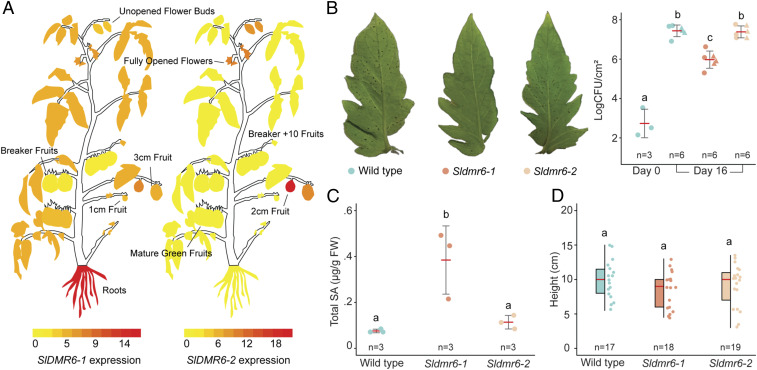
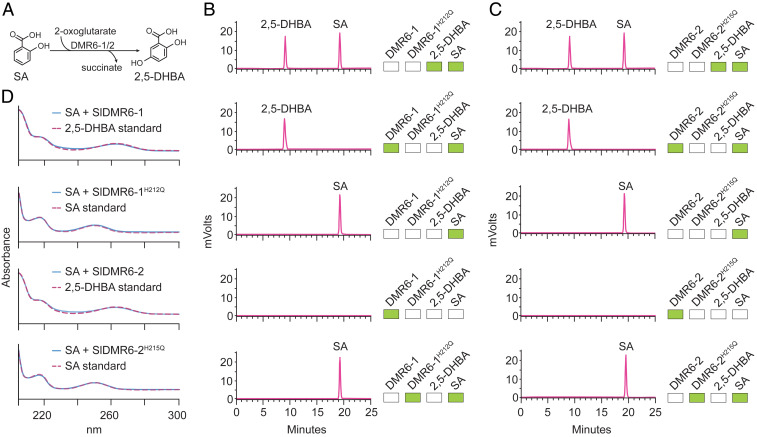
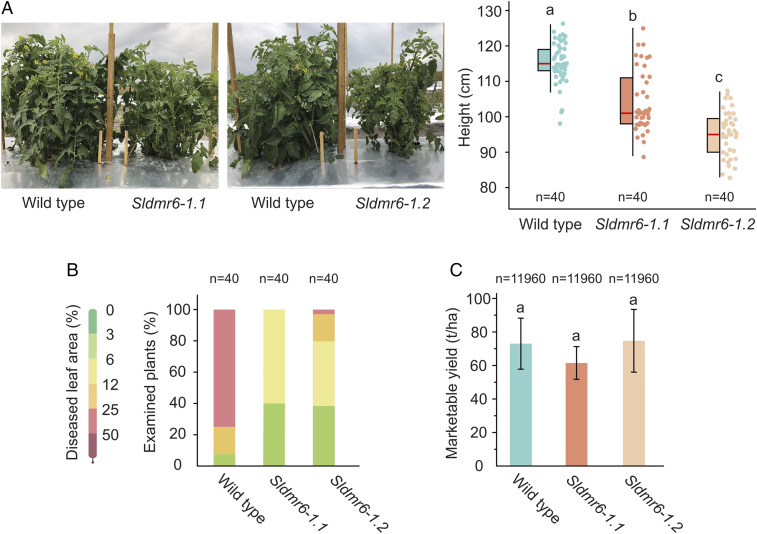
Plant diseases are among the major causes of crop yield losses around the world. To confer disease resistance, conventional breeding relies on the deployment of single resistance (R) genes. However, this strategy has been easily overcome by constantly evolving pathogens. Disabling susceptibility (S) genes is a promising alternative to R genes in breeding programs, as it usually offers durable and broad-spectrum disease resistance. In Arabidopsis, the S gene DMR6 (AtDMR6) encodes an enzyme identified as a susceptibility factor to bacterial and oomycete pathogens. Here, we present a model-to-crop translational work in which we characterize two AtDMR6 orthologs in tomato, SlDMR6-1 and SlDMR6-2. We show that SlDMR6-1, but not SlDMR6-2, is up-regulated by pathogen infection. In agreement, Sldmr6-1 mutants display enhanced resistance against different classes of pathogens, such as bacteria, oomycete, and fungi. Notably, disease resistance correlates with increased salicylic acid (SA) levels and transcriptional activation of immune responses. Furthermore, we demonstrate that SlDMR6-1 and SlDMR6-2 display SA-5 hydroxylase activity, thus contributing to the elucidation of the enzymatic function of DMR6. We then propose that SlDMR6 duplication in tomato resulted in subsequent subfunctionalization, in which SlDMR6-2 specialized in balancing SA levels in flowers/fruits, while SlDMR6-1 conserved the ability to fine-tune SA levels during pathogen infection of the plant vegetative tissues. Overall, this work not only corroborates a mechanism underlying SA homeostasis in plants, but also presents a promising strategy for engineering broad-spectrum and durable disease resistance in crops.
Keywords: CRISPR/Cas9 technology; DMR6; crop engineering; disease resistance; salicylic acid.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Savary S., et al., The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 3, 430–439 (2019). - PubMed
-
- Fuchs M., Pyramiding resistance-conferring gene sequences in crops. Curr. Opin. Virol. 26, 36–42 (2017). - PubMed
-
- Mundt C. C., Pyramiding for resistance durability: Theory and practice. Phytopathology 108, 792–802 (2018). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
