

Protein arginine methyltransferase 8 modulates mitochondrial bioenergetics and neuroinflammation after hypoxic stress
- PMID: 34216036
- PMCID: PMC8595568
- DOI: 10.1111/jnc.15462
Protein arginine methyltransferase 8 modulates mitochondrial bioenergetics and neuroinflammation after hypoxic stress
Abstract
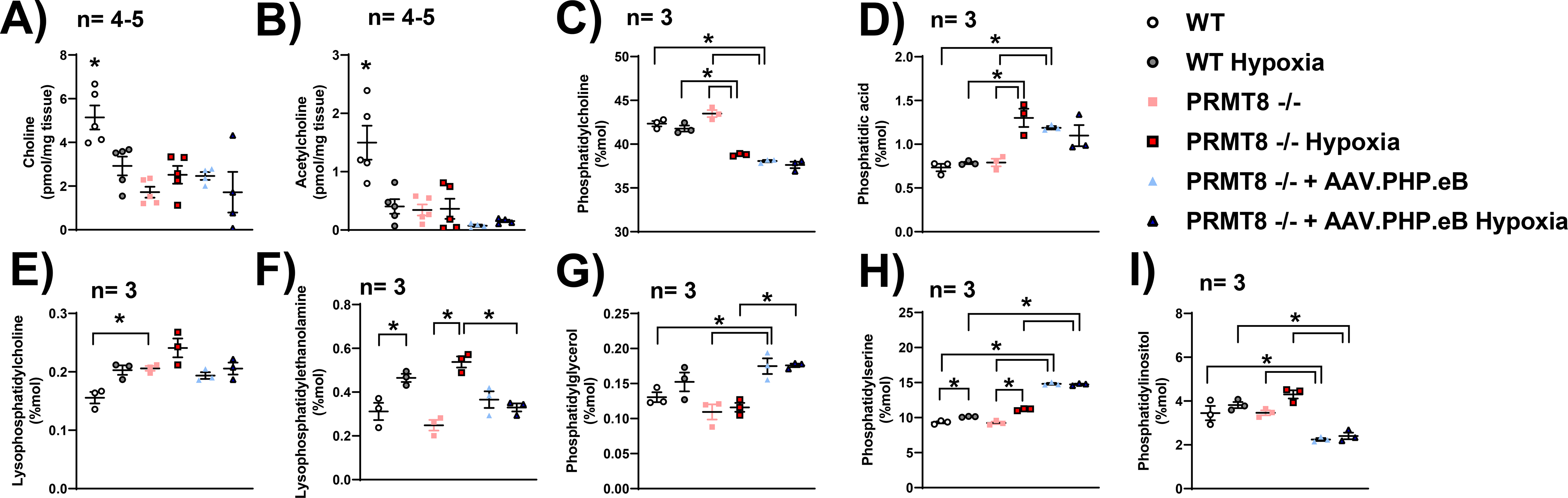
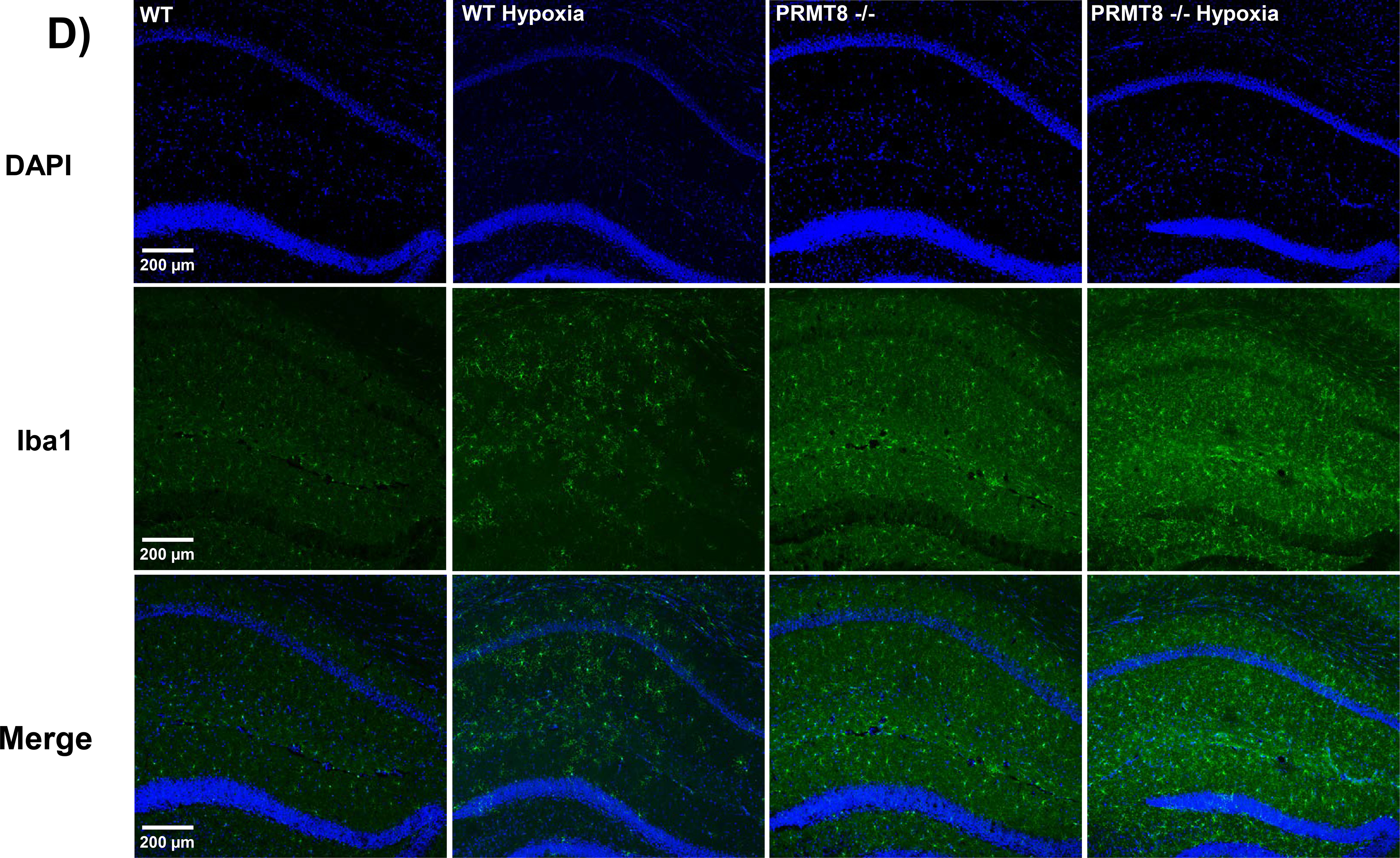
Protein arginine methyltransferases (PRMTs) are a family of enzymes involved in gene regulation and protein/histone modifications. PRMT8 is primarily expressed in the central nervous system, specifically within the cellular membrane and synaptic vesicles. Recently, PRMT8 has been described to play key roles in neuronal signaling such as a regulator of dendritic arborization, synaptic function and maturation, and neuronal differentiation and plasticity. Here, we examined the role of PRMT8 in response to hypoxia-induced stress in brain metabolism. Our results from liquid chromatography mass spectrometry, mitochondrial oxygen consumption rate, and protein analyses indicate that PRMT8(-/-) knockout mice presented with altered membrane phospholipid composition, decreased mitochondrial stress capacity, and increased neuroinflammatory markers, such as tumor necrosis factor alpha and ionized calcium binding adaptor molecule 1 (Iba1, a specific marker for microglia/macrophage activation) after hypoxic stress. Furthermore, adenovirus-based overexpression of PRMT8 reversed the changes in membrane phospholipid composition, mitochondrial stress capacity, and neuroinflammatory markers. Together, our findings establish PRMT8 as an important regulatory component of membrane phospholipid composition, short-term memory function, mitochondrial function, and neuroinflammation in response to hypoxic stress.
Keywords: hypoxic stress; mitochondrial function; neuroinflammation; phospholipids; protein arginine methyltransferase.
© 2021 International Society for Neurochemistry.
Conflict of interest statement
Conflict of Interest Disclosure
No conflicts of interest, financial or otherwise, are declared by the author(s).
--Human subjects --
Involves human subjects:
If yes: Informed consent & ethics approval achieved:
=> if yes, please ensure that the info “Informed consent was achieved for all subjects, and the experiments were approved by the local ethics committee.” is included in the Methods.
ARRIVE guidelines have been followed:
Yes
=> if it is a Review or Editorial, skip complete sentence => if No, include a statement in the “Conflict of interest disclosure” section: “ARRIVE guidelines were not followed for the following reason:
“
(edit phrasing to form a complete sentence as necessary).
=> if Yes, insert in the “Conflict of interest disclosure” section:
“All experiments were conducted in compliance with the ARRIVE guidelines.” unless it is a Review or Editorial
Figures








References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
