

Predominantly symplastic phloem unloading of photosynthates maintains efficient starch accumulation in the cassava storage roots (Manihot esculenta Crantz)
- PMID: 34217217
- PMCID: PMC8254309
- DOI: 10.1186/s12870-021-03088-1
Predominantly symplastic phloem unloading of photosynthates maintains efficient starch accumulation in the cassava storage roots (Manihot esculenta Crantz)
Abstract
Background: Cassava (Manihot esculenta Crantz) efficiently accumulates starch in its storage roots. However, how photosynthates are transported from the leaves to the phloem (especially how they are unloaded into parenchymal cells of storage roots) remains unclear.
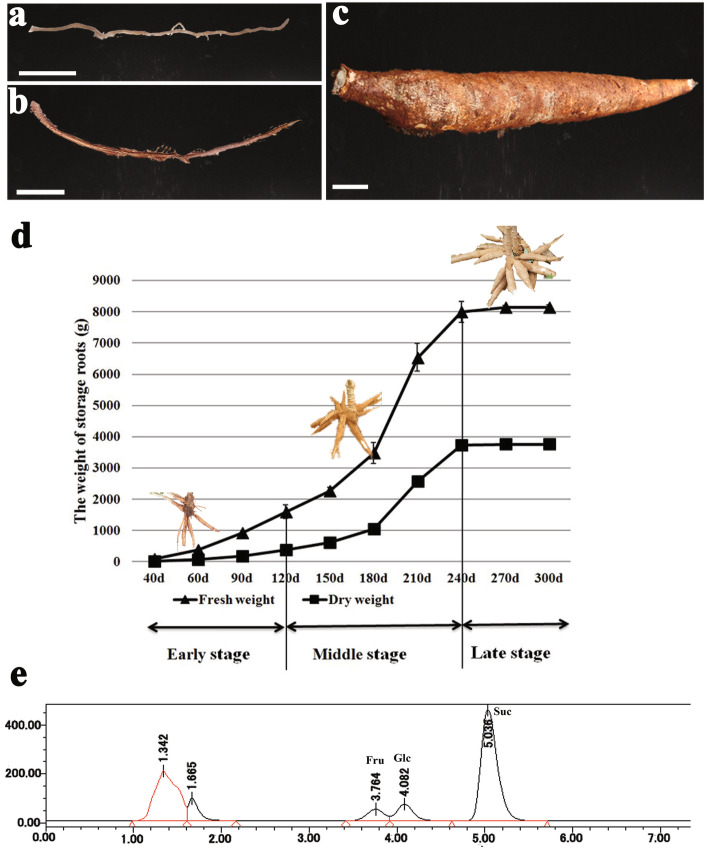
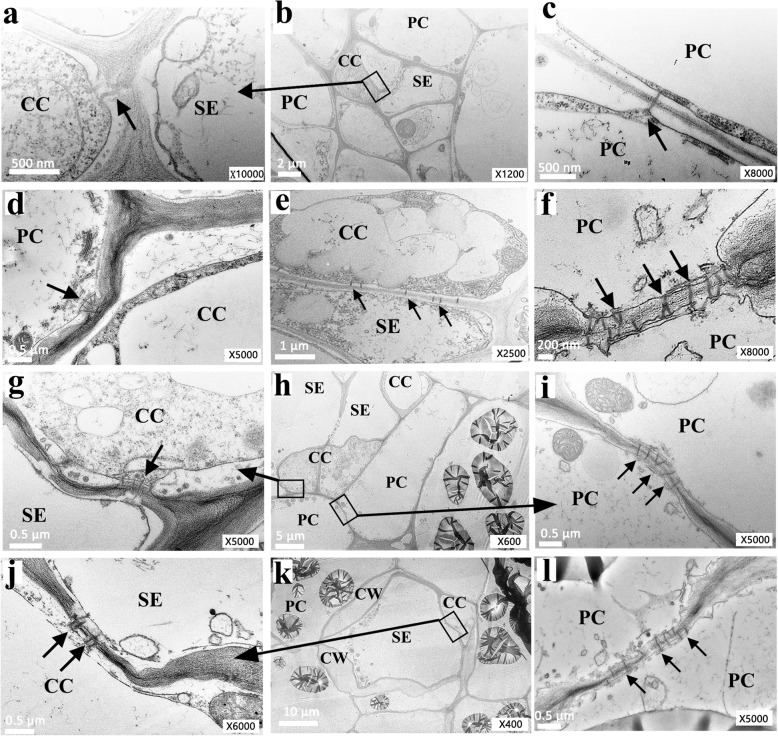
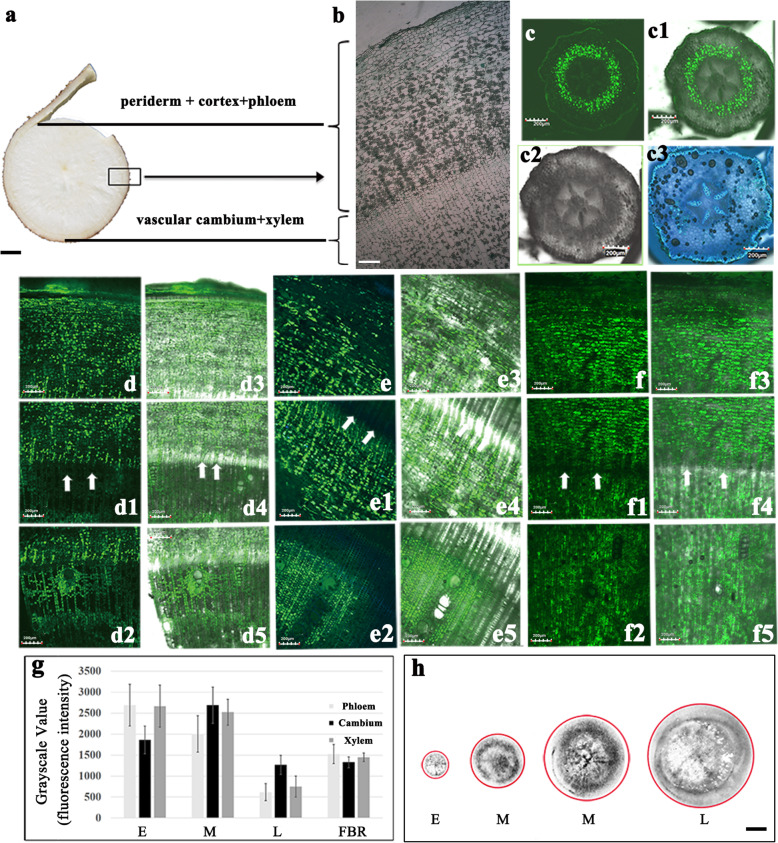
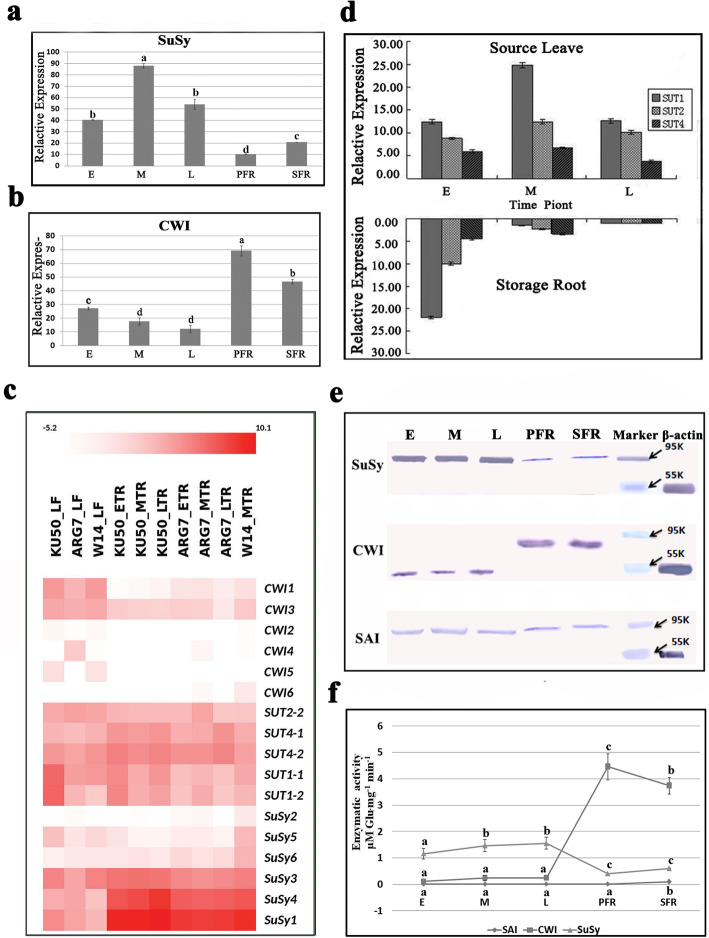
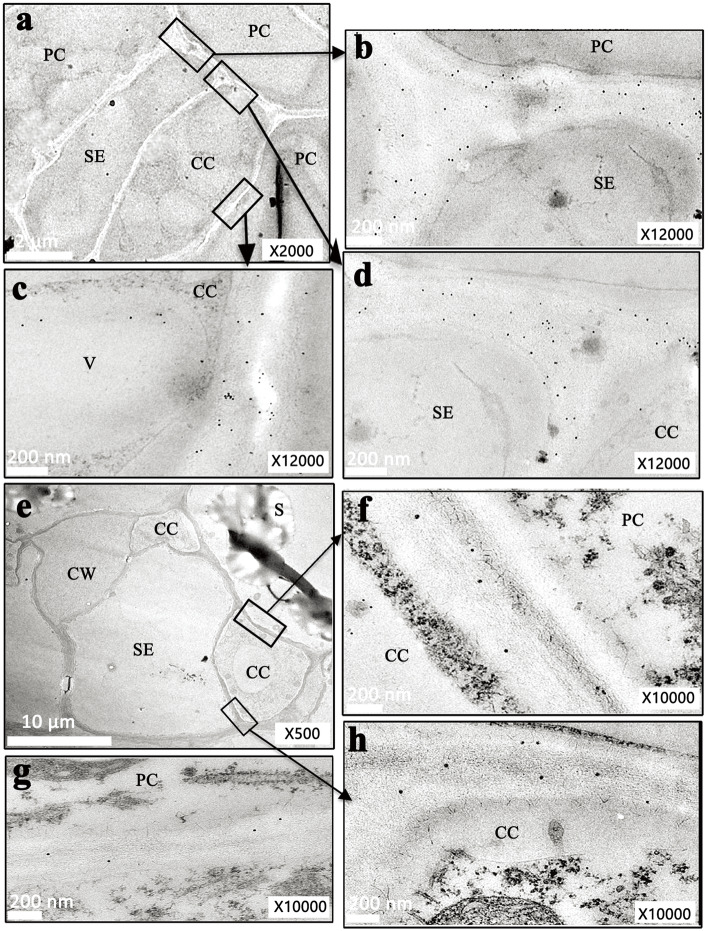
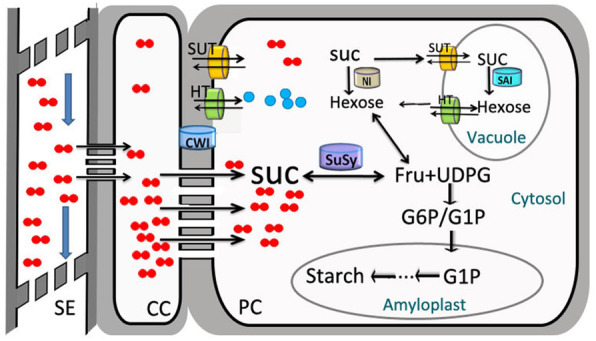
Results: Here, we investigated the sucrose unloading pattern and its impact on cassava storage root development using microstructural and physiological analyses, namely, carboxyfluorescein (CF) and C14 isotope tracing. The expression profiling of genes involved in symplastic and apoplastic transport was performed, which included enzyme activity, protein gel blot analysis, and transcriptome sequencing analyses. These finding showed that carbohydrates are transported mainly in the form of sucrose, and more than 54.6% was present in the stem phloem. Sucrose was predominantly unloaded symplastically from the phloem into storage roots; in addition, there was a shift from apoplastic to symplastic unloading accompanied by the onset of root swelling. Statistical data on the microstructures indicated an enrichment of plasmodesmata within sieve, companion, and parenchyma cells in the developing storage roots of a cultivar but not in a wild ancestor. Tracing tests with CF verified the existence of a symplastic channel, and [14C] Suc demonstrated that sucrose could rapidly diffuse into root parenchyma cells from phloem cells. The relatively high expression of genes encoding sucrose synthase and associated proteins appeared in the middle and late stages of storage roots but not in primary fibrous roots, or secondary fibrous roots. The inverse expression pattern of sucrose transporters, cell wall acid invertase, and soluble acid invertase in these corresponding organs supported the presence of a symplastic sucrose unloading pathway. The transcription profile of genes involved in symplastic unloading and their significantly positive correlation with the starch yield at the population level confirmed that symplastic sucrose transport is vitally important in the development of cassava storage roots.
Conclusions: In this study, we revealed that the cassava storage root phloem sucrose unloading pattern was predominantly a symplastic unloading pattern. This pattern is essential for efficient starch accumulation in high-yielding varieties compared with low-yielding wild ancestors.
Keywords: Carbohydrates; Cassava; Starch yield; Storage root; Symplastic unloading.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Frommer WB, Sonnewald U. Molecular analysis of carbon partitioning in solanaceous species. J Exp Bot. 1995;46(6):587–607. doi: 10.1093/jxb/46.6.587. - DOI
-
- Ishimaru T, Hirose T, Matsuda T, Goto A, Takahashi K, Sasaki H, Terao T, Ishii R, Ohsugi R, Yamagishi T. Expression patterns of genes encoding carbohydrate-metabolizing enzymes and their relationship to grain filling in rice (Oryza sativa L.): comparison of caryopses located at different positions in a panicle. Plant Cell Physiol. 2005;46(4):620–628. doi: 10.1093/pcp/pci066. - DOI - PubMed
-
- Oparka KJ, Roberts AG, Boevink P, Cruz SS, Roberts I, Pradel KS, Imlau A, Otlizky GK, Kotlizky G, Sauer N, et al. Simple, but not branched, plasmodesmata allow the nonspecific trafficking of proteins in developing tobacco leaves. Cell. 1999;97(6):743–754. doi: 10.1016/S0092-8674(00)80786-2. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
