

ALKBH3 partner ASCC3 mediates P-body formation and selective clearance of MMS-induced 1-methyladenosine and 3-methylcytosine from mRNA
- PMID: 34217309
- PMCID: PMC8254245
- DOI: 10.1186/s12967-021-02948-6
ALKBH3 partner ASCC3 mediates P-body formation and selective clearance of MMS-induced 1-methyladenosine and 3-methylcytosine from mRNA
Abstract
Background: Reversible enzymatic methylation of mammalian mRNA is widespread and serves crucial regulatory functions, but little is known to what degree chemical alkylators mediate overlapping modifications and whether cells distinguish aberrant from canonical methylations.
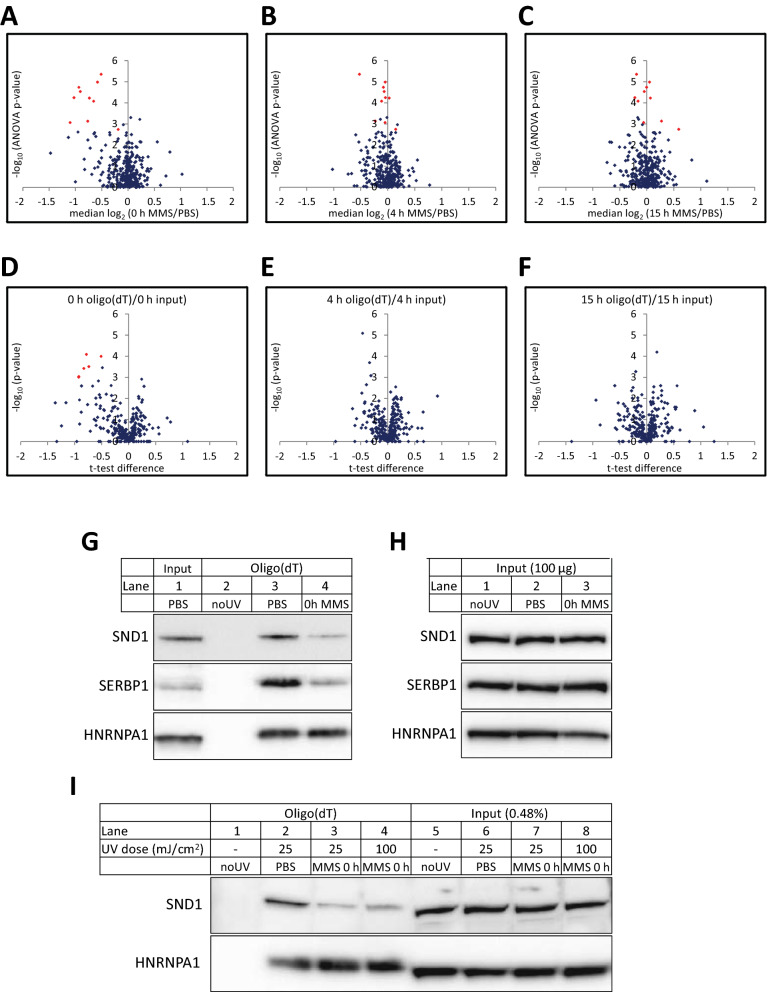
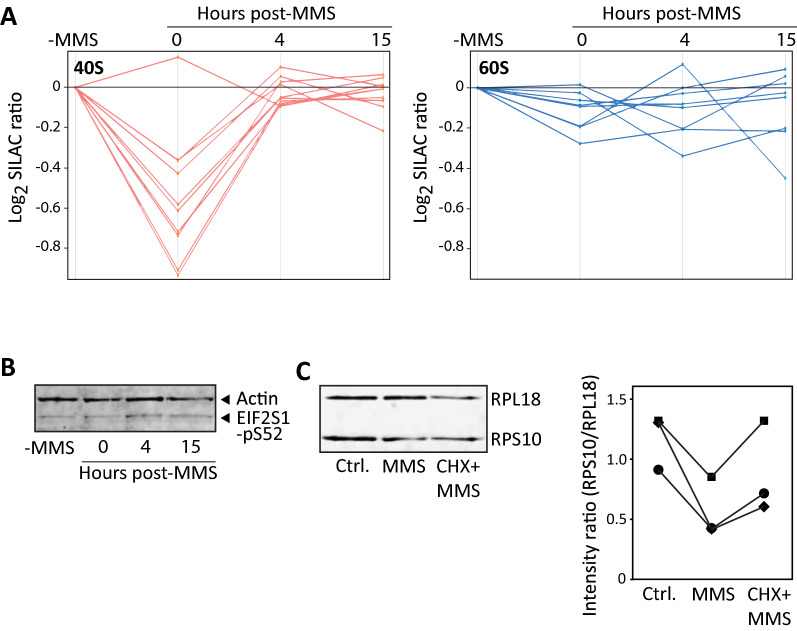
Methods: Here we use quantitative mass spectrometry to determine the fate of chemically induced methylbases in the mRNA of human cells. Concomitant alteration in the mRNA binding proteome was analyzed by SILAC mass spectrometry.
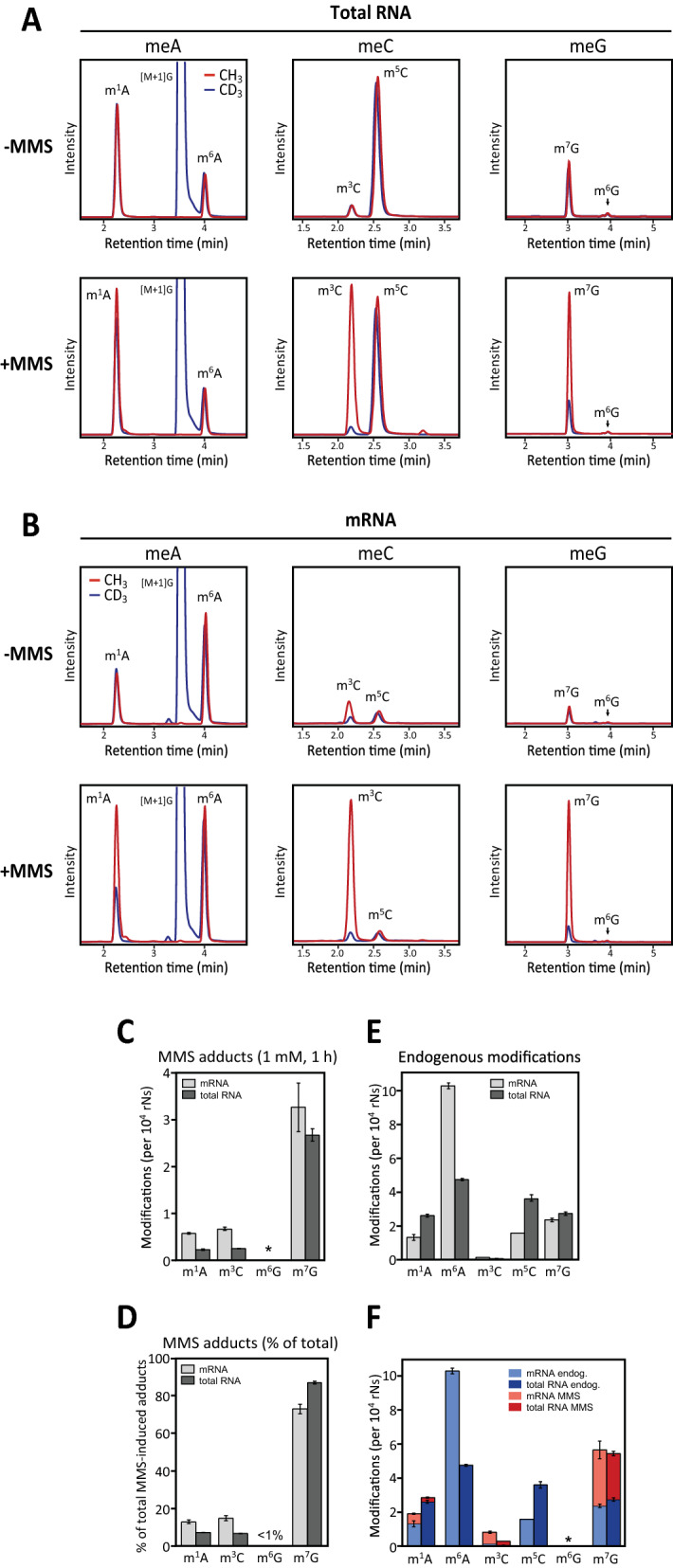
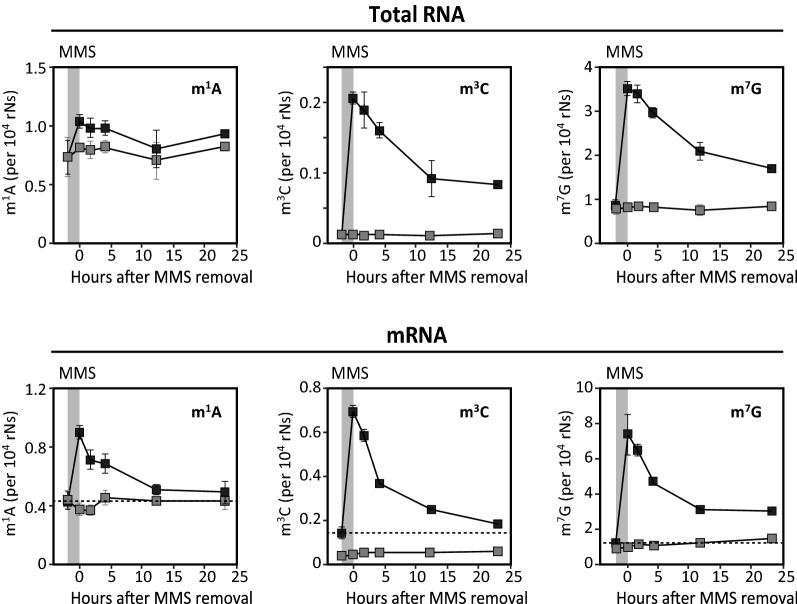
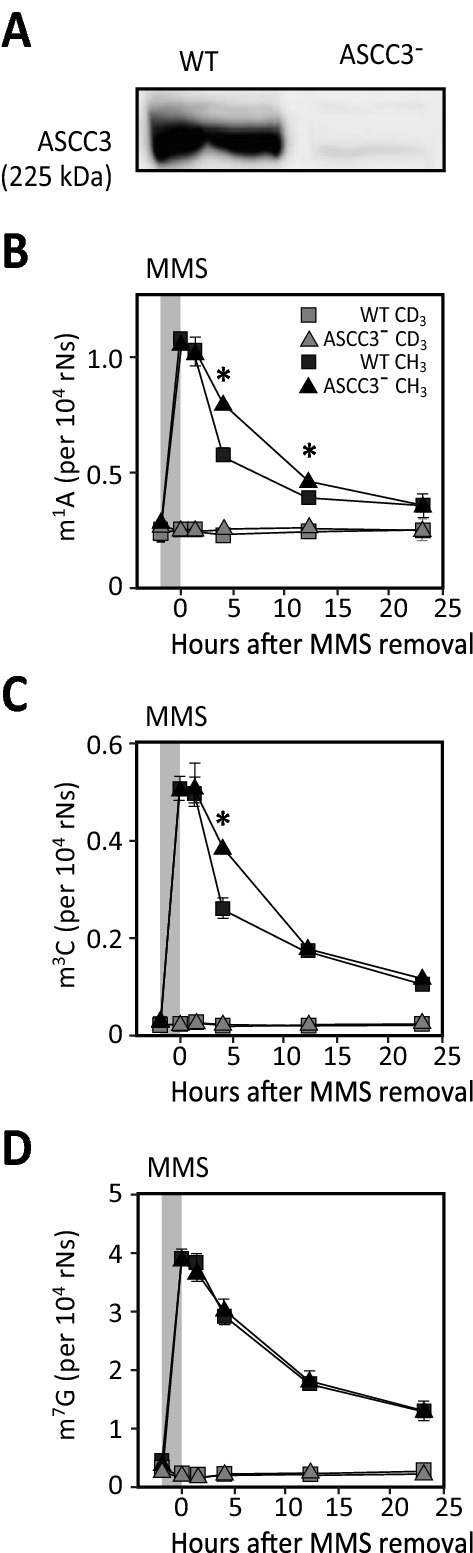
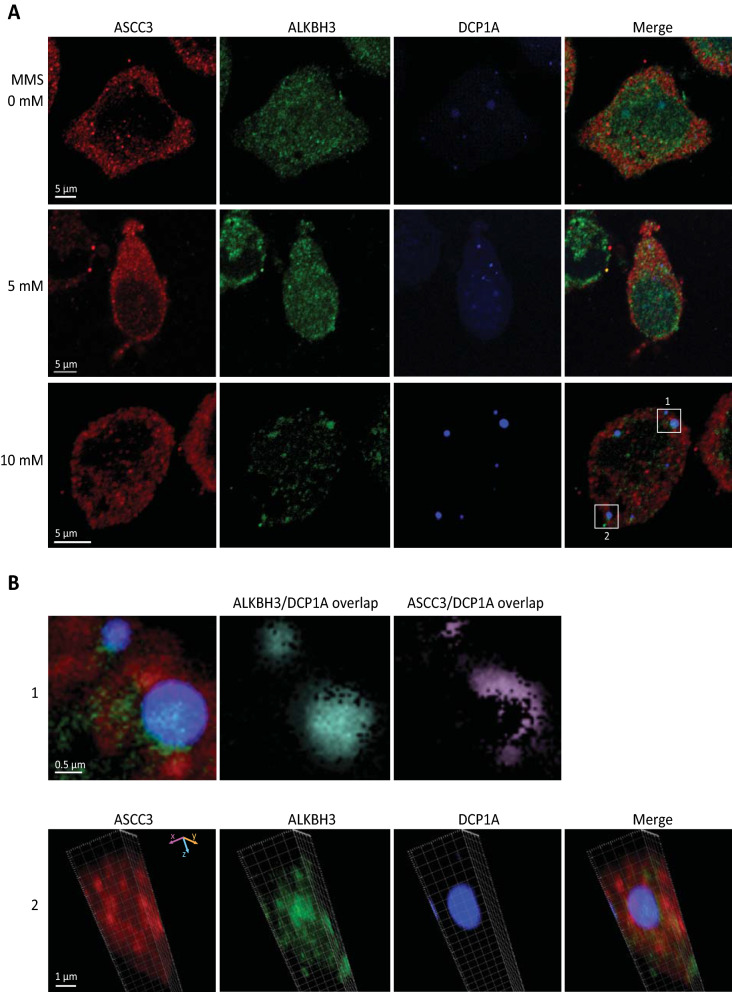
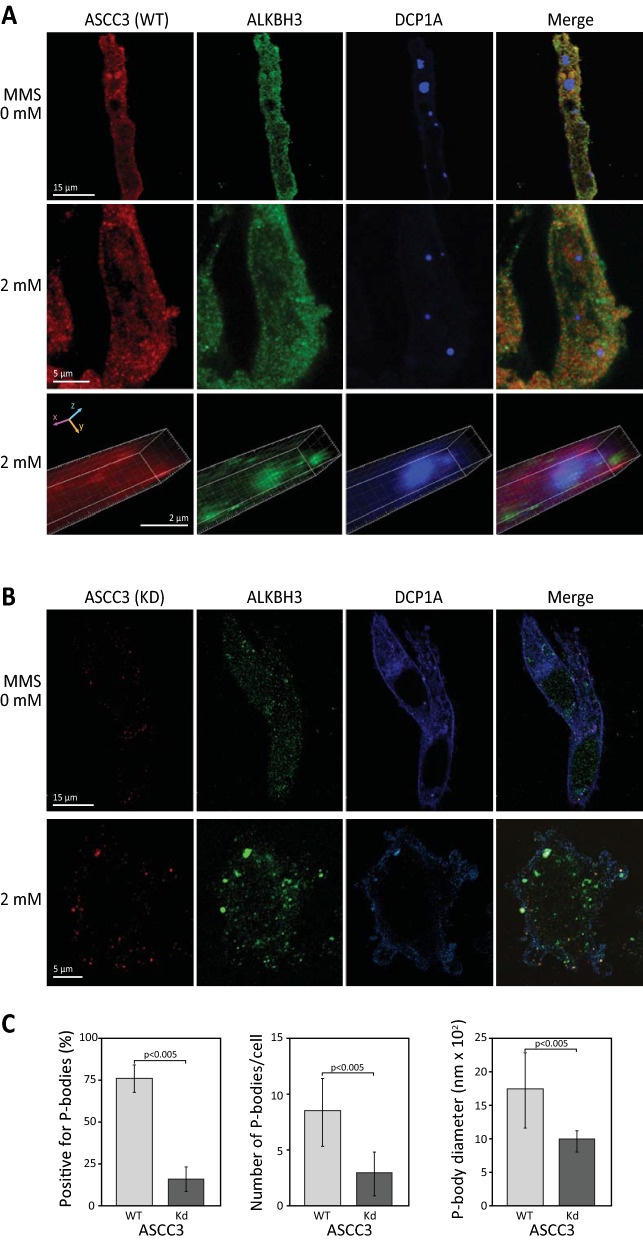
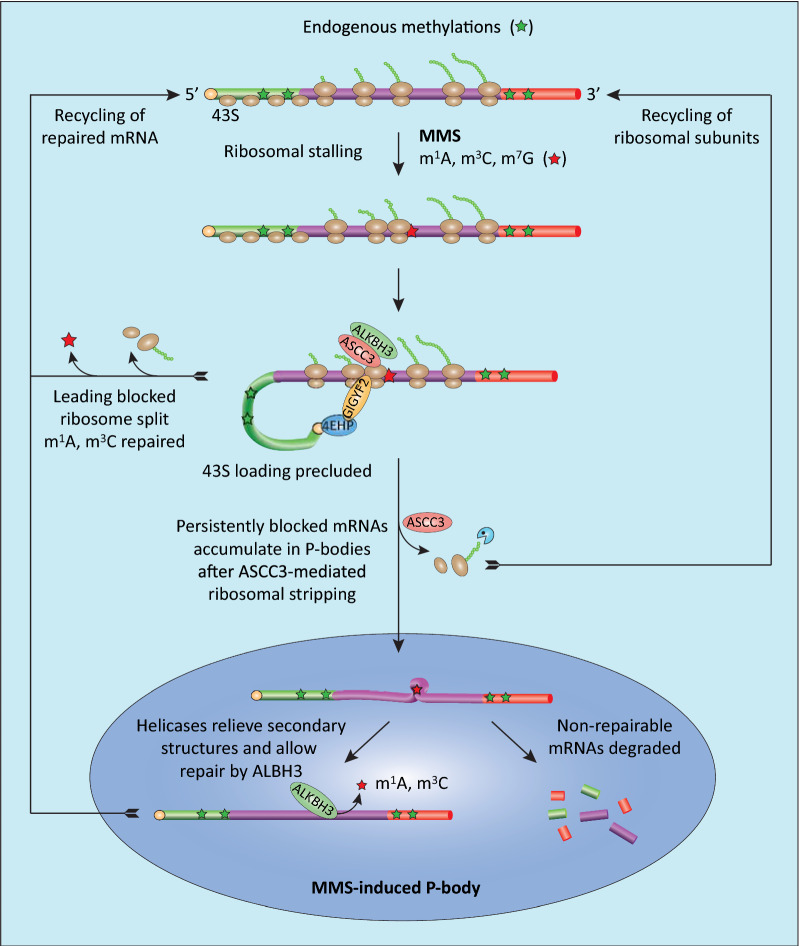
Results: MMS induced prominent direct mRNA methylations that were chemically identical to endogenous methylbases. Transient loss of 40S ribosomal proteins from isolated mRNA suggests that aberrant methylbases mediate arrested translational initiation and potentially also no-go decay of the affected mRNA. Four proteins (ASCC3, YTHDC2, TRIM25 and GEMIN5) displayed increased mRNA binding after MMS treatment. ASCC3 is a binding partner of the DNA/RNA demethylase ALKBH3 and was recently shown to promote disassembly of collided ribosomes as part of the ribosome quality control (RQC) trigger complex. We find that ASCC3-deficient cells display delayed removal of MMS-induced 1-methyladenosine (m1A) and 3-methylcytosine (m3C) from mRNA and impaired formation of MMS-induced P-bodies.
Conclusions: Our findings conform to a model in which ASCC3-mediated disassembly of collided ribosomes allows demethylation of aberrant m1A and m3C by ALKBH3. Our findings constitute first evidence of selective sanitation of aberrant mRNA methylbases over their endogenous counterparts and warrant further studies on RNA-mediated effects of chemical alkylators commonly used in the clinic.
Keywords: 1-Methyladenosine; 3-Methylcytosine; 7-Methylguanosine; ALKBH3; ASCC3; Alkylating agents; Epitranscriptome; No-go decay; P-bodies; Ribosome quality control.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
