

Glutamate Dehydrogenase Is Important for Ammonia Fixation and Amino Acid Homeostasis in Brain During Hyperammonemia
- PMID: 34220417
- PMCID: PMC8244593
- DOI: 10.3389/fnins.2021.646291
Glutamate Dehydrogenase Is Important for Ammonia Fixation and Amino Acid Homeostasis in Brain During Hyperammonemia
Abstract
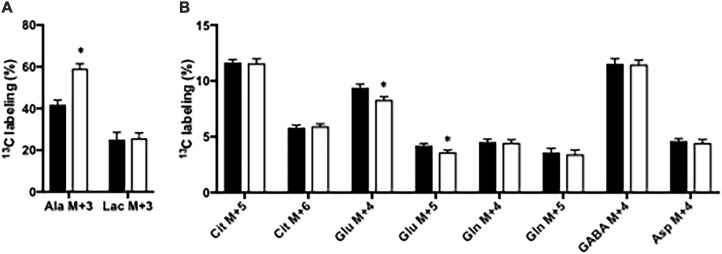
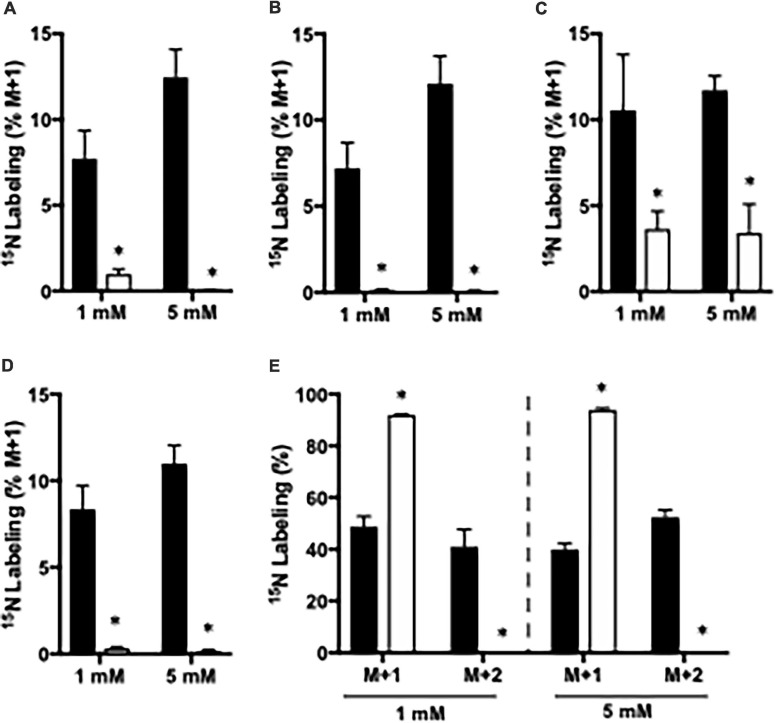
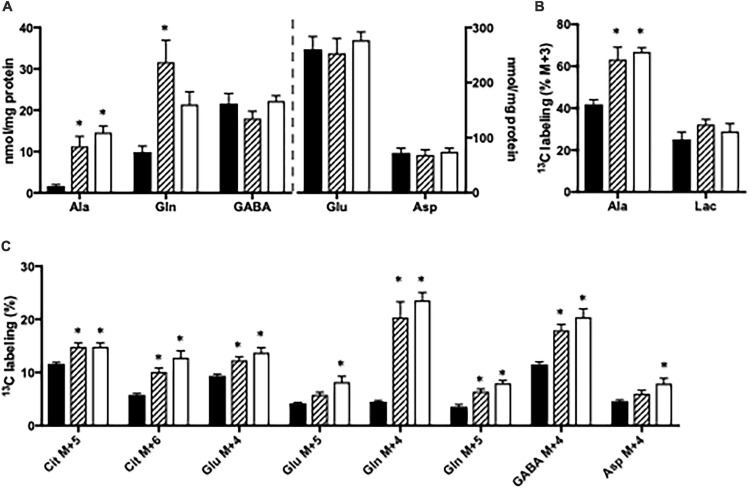
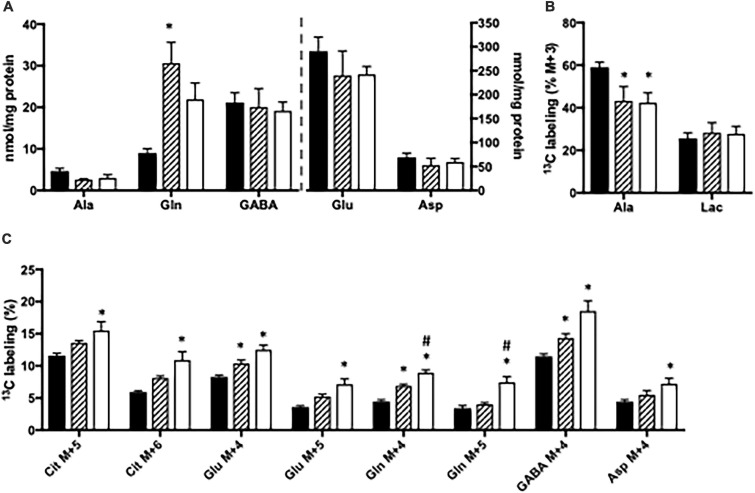
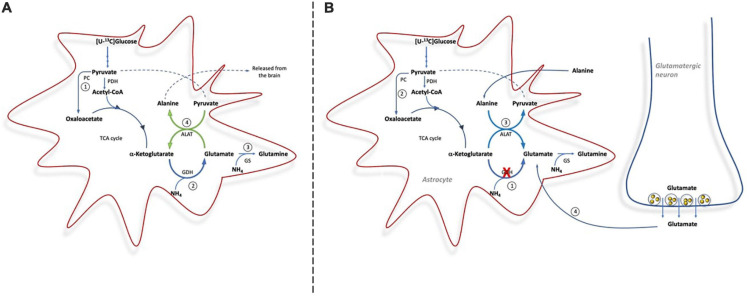
Impaired liver function may lead to hyperammonemia and risk for hepatic encephalopathy. In brain, detoxification of ammonia is mediated mainly by glutamine synthetase (GS) in astrocytes. This requires a continuous de novo synthesis of glutamate, likely involving the action of both pyruvate carboxylase (PC) and glutamate dehydrogenase (GDH). An increased PC activity upon ammonia exposure and the importance of PC activity for glutamine synthesis has previously been demonstrated while the importance of GDH for generation of glutamate as precursor for glutamine synthesis has received little attention. We therefore investigated the functional importance of GDH for brain metabolism during hyperammonemia. To this end, brain slices were acutely isolated from transgenic CNS-specific GDH null or litter mate control mice and incubated in aCSF containing [U-13C]glucose in the absence or presence of 1 or 5 mM ammonia. In another set of experiments, brain slices were incubated in aCSF containing 1 or 5 mM 15N-labeled NH4Cl and 5 mM unlabeled glucose. Tissue extracts were analyzed for isotopic labeling in metabolites and for total amounts of amino acids. As a novel finding, we reveal a central importance of GDH function for cerebral ammonia fixation and as a prerequisite for de novo synthesis of glutamate and glutamine during hyperammonemia. Moreover, we demonstrated an important role of the concerted action of GDH and alanine aminotransferase in hyperammonemia; the products alanine and α-ketoglutarate serve as an ammonia sink and as a substrate for ammonia fixation via GDH, respectively. The role of this mechanism in human hyperammonemic states remains to be studied.
Keywords: alanine; brain; glutamate; glutamate dehydogenase; glutamine; hyperammonemia; pyruvate carboxylase (PC).
Copyright © 2021 Voss, Arildsen, Nissen, Waagepetersen, Schousboe, Maechler, Ott, Vilstrup and Walls.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Inhibition of glutamine synthesis induces glutamate dehydrogenase-dependent ammonia fixation into alanine in co-cultures of astrocytes and neurons.Neurochem Int. 2011 Sep;59(4):482-8. doi: 10.1016/j.neuint.2011.03.008. Epub 2011 Mar 21. Neurochem Int. 2011. PMID: 21397649
-
Brain alanine formation as an ammonia-scavenging pathway during hyperammonemia: effects of glutamine synthetase inhibition in rats and astrocyte-neuron co-cultures.J Cereb Blood Flow Metab. 2013 Aug;33(8):1235-41. doi: 10.1038/jcbfm.2013.73. Epub 2013 May 15. J Cereb Blood Flow Metab. 2013. PMID: 23673435 Free PMC article.
-
Central Role of Glutamate Metabolism in the Maintenance of Nitrogen Homeostasis in Normal and Hyperammonemic Brain.Biomolecules. 2016 Mar 26;6(2):16. doi: 10.3390/biom6020016. Biomolecules. 2016. PMID: 27023624 Free PMC article. Review.
-
Glutamine synthetase in brain: effect of ammonia.Neurochem Int. 2002 Aug-Sep;41(2-3):123-42. doi: 10.1016/s0197-0186(02)00033-5. Neurochem Int. 2002. PMID: 12020613 Review.
-
Role of pyruvate carboxylase in facilitation of synthesis of glutamate and glutamine in cultured astrocytes.J Neurochem. 1997 Dec;69(6):2312-25. doi: 10.1046/j.1471-4159.1997.69062312.x. J Neurochem. 1997. PMID: 9375662
Cited by
-
Evolution of Glutamate Metabolism via GLUD2 Enhances Lactate-Dependent Synaptic Plasticity and Complex Cognition.Int J Mol Sci. 2024 May 13;25(10):5297. doi: 10.3390/ijms25105297. Int J Mol Sci. 2024. PMID: 38791334 Free PMC article. Review.
-
Fueling Brain Inhibition: Integrating GABAergic Neurotransmission and Energy Metabolism.Neurochem Res. 2025 Apr 7;50(2):136. doi: 10.1007/s11064-025-04384-0. Neurochem Res. 2025. PMID: 40189668 Review.
-
Unveiling metabolic integration in psyllids and their nutritional endosymbionts through comparative transcriptomics analysis.iScience. 2023 Sep 15;26(10):107930. doi: 10.1016/j.isci.2023.107930. eCollection 2023 Oct 20. iScience. 2023. PMID: 37810228 Free PMC article.
-
Integrated multi‑omics analysis of liver metabolic dysregulation in ACE2 knockout mice.Int J Mol Med. 2025 Sep;56(3):134. doi: 10.3892/ijmm.2025.5575. Epub 2025 Jul 4. Int J Mol Med. 2025. PMID: 40613222 Free PMC article.
-
The Relationship between Plasma Alpha-1-Antitrypsin Polymers and Lung or Liver Function in ZZ Alpha-1-Antitrypsin-Deficient Patients.Biomolecules. 2022 Feb 28;12(3):380. doi: 10.3390/biom12030380. Biomolecules. 2022. PMID: 35327571 Free PMC article.
References
-
- Berl S., Takagaki G., Clarke D. D., Waelsch H. (1962). Carbon dioxide fixation in the brain. J. Biol. Chem. 237 2570–2573. - PubMed
-
- Biemann K. (1962). Mass Spectrometry, Organic Chemistry Applications. New York: McGraw, 223–227.
-
- Bosman D. K., Deutz N. E., De Graaf A. A., vd Hulst R. W., Van Eijk H. M., Bovee W. M., et al. (1990). Changes in brain metabolism during hyperammonemia and acute liver failure: results of a comparative 1H-NMR spectroscopy and biochemical investigation. Hepatology 12 281–290. 10.1002/hep.1840120215 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials