

The evolutionary dynamics of viruses: virion release strategies, time delays and fitness minima
- PMID: 34221452
- PMCID: PMC8242231
- DOI: 10.1093/ve/veab039
The evolutionary dynamics of viruses: virion release strategies, time delays and fitness minima
Abstract
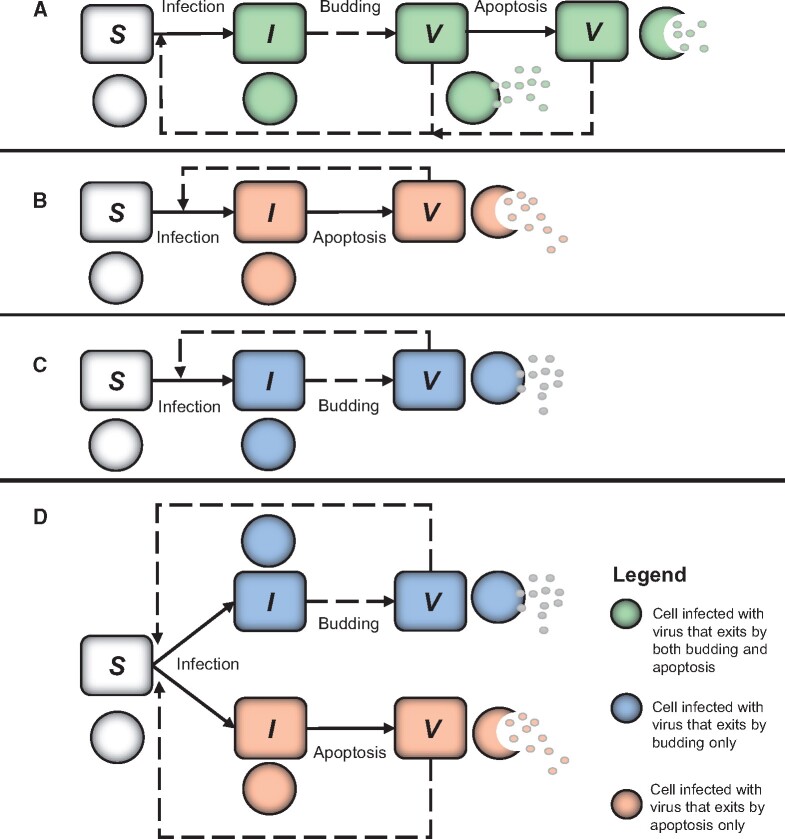
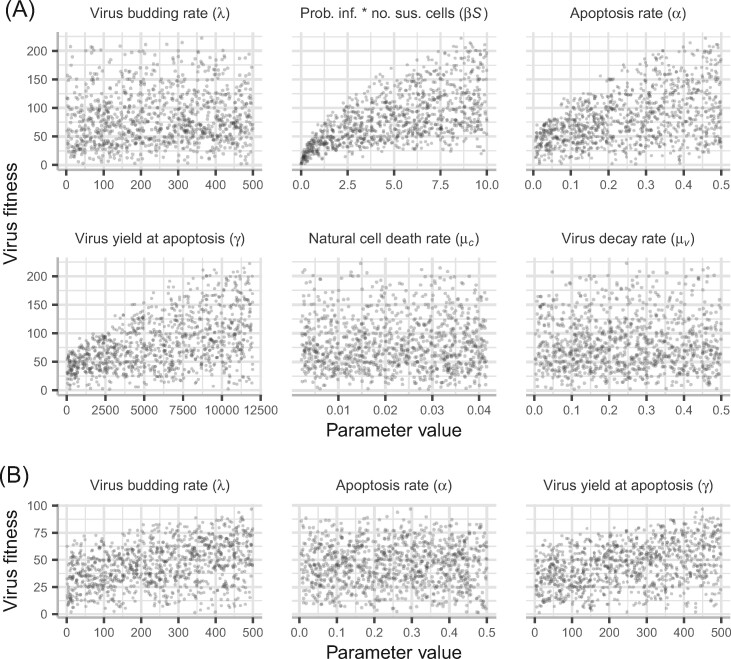
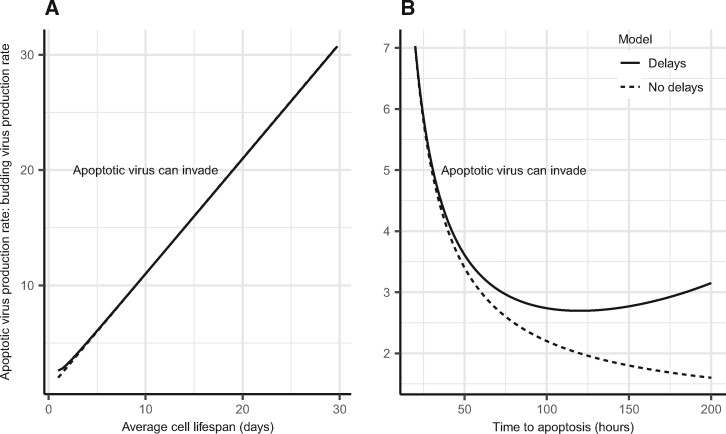
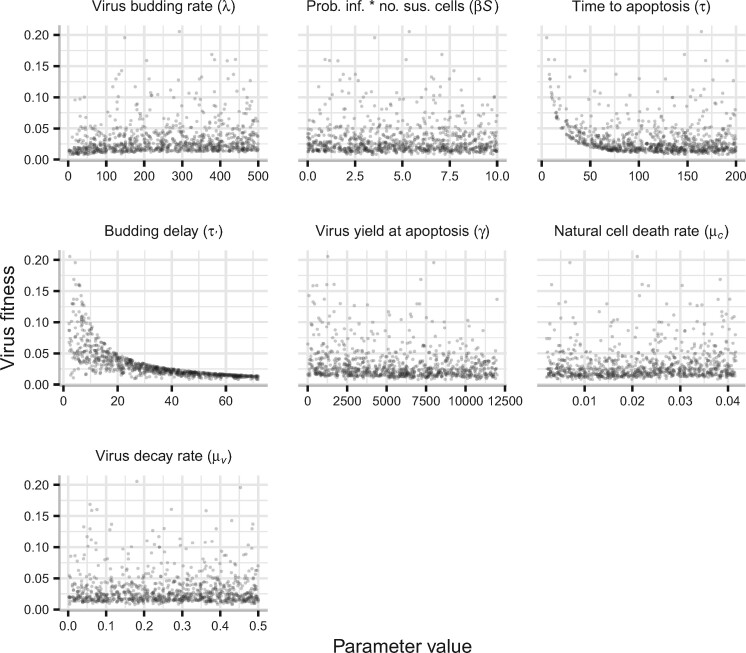
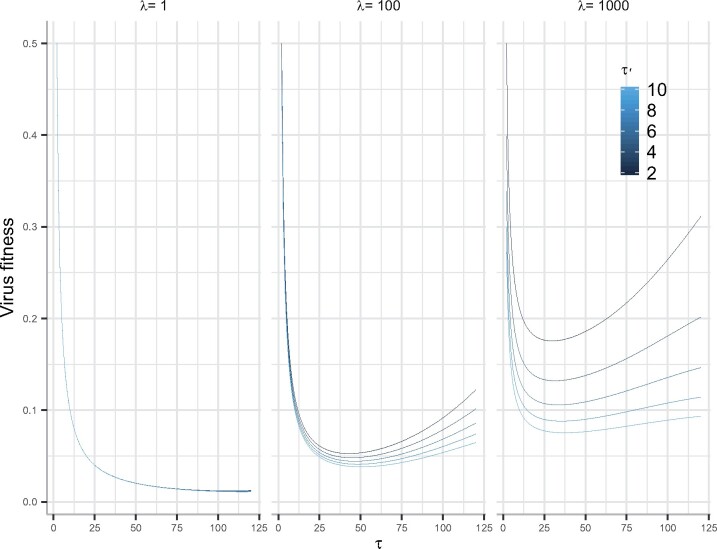
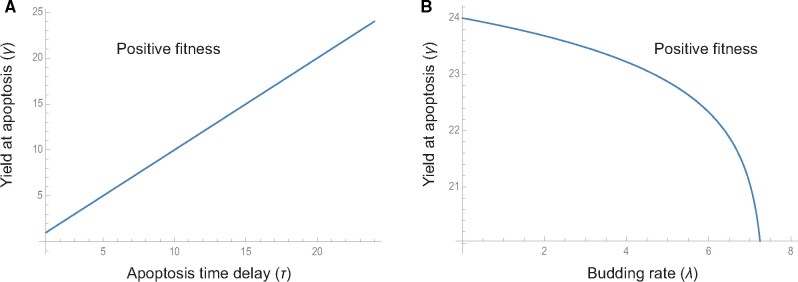
Viruses exhibit a diverse array of strategies for infecting host cells and for virion release after replication. Cell exit strategies generally involve either budding from the cell membrane or killing the host cell. The conditions under which either is at a selective advantage is a key question in the evolutionary theory of viruses, with the outcome having potentially important impacts on the course of infection and pathogenicity. Although a plethora of external factors will influence the fitness of either strategy; here, we focus just on the effects of the physical properties of the system. We develop theoretical approaches to assess the effects of the time delays between initial infection and virion release. We show that the length of the delay before apoptosis is an important trait in virus evolutionary dynamics. Our results show that for a fixed time to apoptosis, intermediate delays lead to virus fitness that is lower than short times to apoptosis - leading to an apoptotic strategy - and long times to apoptosis - leading to a budding strategy at the between-cell level. At fitness minima, selection is expected to be disruptive and the potential for adaptive radiation in virus strategies is feasible. Hence, the physical properties of the system are sufficient to explain the existence of both budding and virus-induced apoptosis. The fitness functions presented here provide a formal basis for further work focusing on the evolutionary implications of trade-offs between time delays, intracellular replication and resulting mutation rates.
Keywords: invasion fitness; mathematical modeling; virus evolution.
© The Author(s) 2021. Published by Oxford University Press.
Figures
Similar articles
-
Trade-offs between and within scales: environmental persistence and within-host fitness of avian influenza viruses.Proc Biol Sci. 2014 Jul 22;281(1787):20133051. doi: 10.1098/rspb.2013.3051. Proc Biol Sci. 2014. PMID: 24898369 Free PMC article.
-
Variation in within-host replication kinetics among virus genotypes provides evidence of specialist and generalist infection strategies across three salmonid host species.Virus Evol. 2022 Aug 24;8(2):veac079. doi: 10.1093/ve/veac079. eCollection 2022. Virus Evol. 2022. PMID: 36101884 Free PMC article.
-
Viral reproductive strategies: How can lytic viruses be evolutionarily competitive?J Theor Biol. 2007 Dec 21;249(4):766-84. doi: 10.1016/j.jtbi.2007.09.013. Epub 2007 Sep 15. J Theor Biol. 2007. PMID: 17945261
-
Local adaptation of plant viruses: lessons from experimental evolution.Mol Ecol. 2017 Apr;26(7):1711-1719. doi: 10.1111/mec.13836. Epub 2016 Sep 22. Mol Ecol. 2017. PMID: 27612225 Review.
-
How do viruses control mitochondria-mediated apoptosis?Virus Res. 2015 Nov 2;209:45-55. doi: 10.1016/j.virusres.2015.02.026. Epub 2015 Mar 1. Virus Res. 2015. PMID: 25736565 Free PMC article. Review.
Cited by
-
The role of APOBEC3-induced mutations in the differential evolution of monkeypox virus.Virus Evol. 2023 Oct 5;9(2):vead058. doi: 10.1093/ve/vead058. eCollection 2023. Virus Evol. 2023. PMID: 37841642 Free PMC article.
-
The role of APOBEC3-induced mutations in the differential evolution of monkeypox virus.ArXiv [Preprint]. 2023 Aug 7:arXiv:2308.03714v1. ArXiv. 2023. Update in: Virus Evol. 2023 Oct 05;9(2):vead058. doi: 10.1093/ve/vead058. PMID: 37608930 Free PMC article. Updated. Preprint.
-
Incorporating Intracellular Processes in Virus Dynamics Models.Microorganisms. 2024 Apr 30;12(5):900. doi: 10.3390/microorganisms12050900. Microorganisms. 2024. PMID: 38792730 Free PMC article. Review.
-
Mathematical modeling of respiratory viral infection and applications to SARS-CoV-2 progression.Math Methods Appl Sci. 2022 Aug 3:10.1002/mma.8606. doi: 10.1002/mma.8606. Online ahead of print. Math Methods Appl Sci. 2022. PMID: 36247228 Free PMC article.
-
In Silico Evaluation of Paxlovid's Pharmacometrics for SARS-CoV-2: A Multiscale Approach.Viruses. 2022 May 20;14(5):1103. doi: 10.3390/v14051103. Viruses. 2022. PMID: 35632843 Free PMC article.
References
-
- Abedon S. T. (1989) ‘Selection for Bacteriophage Latent Period Length by Bacterial Density: A Theoretical Examination’, Microbial Ecology, 18: 79–88. - PubMed
-
- Alizon S., Michalakis Y. (2015) ‘Adaptive Virulence Evolution: The Good Old Fitness-Based Approach’, Trends in Ecology & Evolution, 30: 248–54. - PubMed
LinkOut - more resources
Full Text Sources
