

Dietary Moutan Cortex Radicis Improves Serum Antioxidant Capacity and Intestinal Immunity and Alters Colonic Microbiota in Weaned Piglets
- PMID: 34222303
- PMCID: PMC8247480
- DOI: 10.3389/fnut.2021.679129
Dietary Moutan Cortex Radicis Improves Serum Antioxidant Capacity and Intestinal Immunity and Alters Colonic Microbiota in Weaned Piglets
Abstract
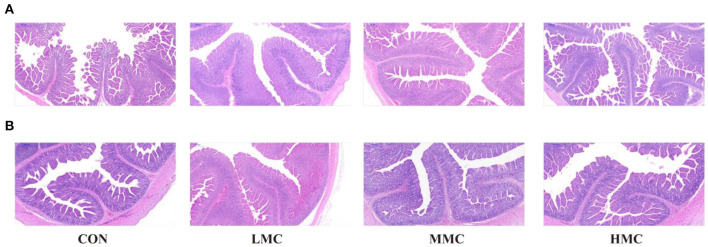
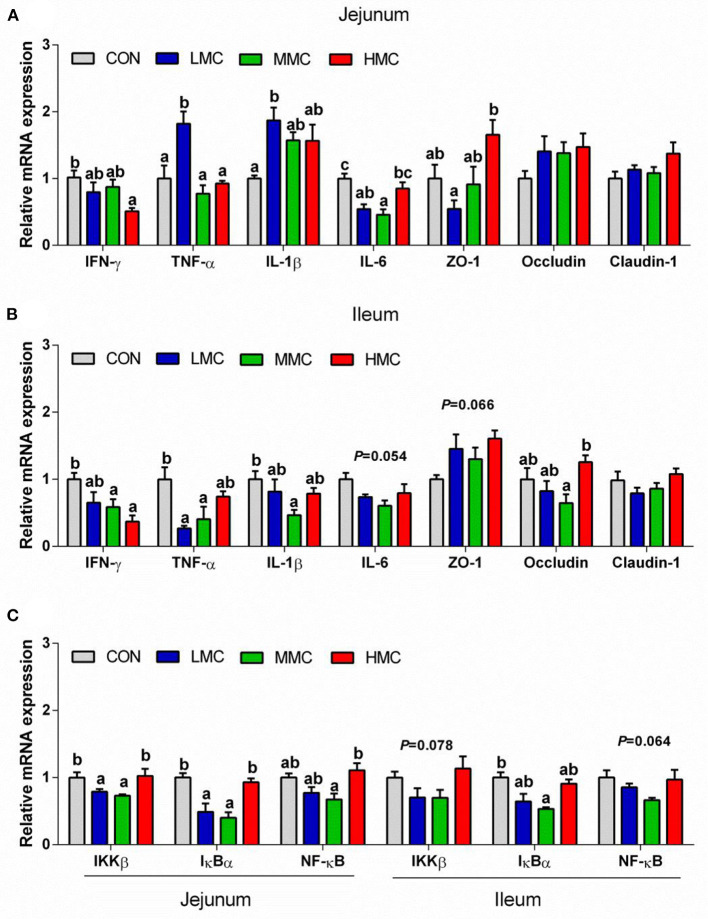
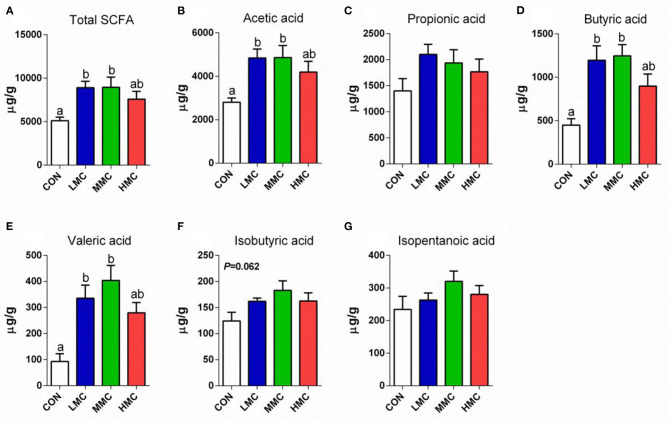
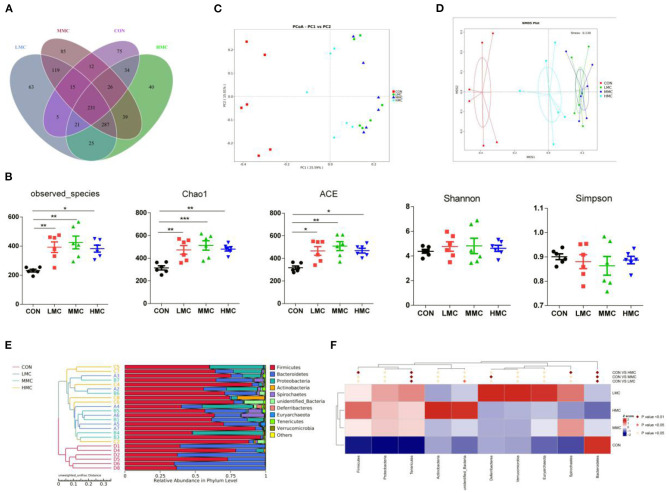
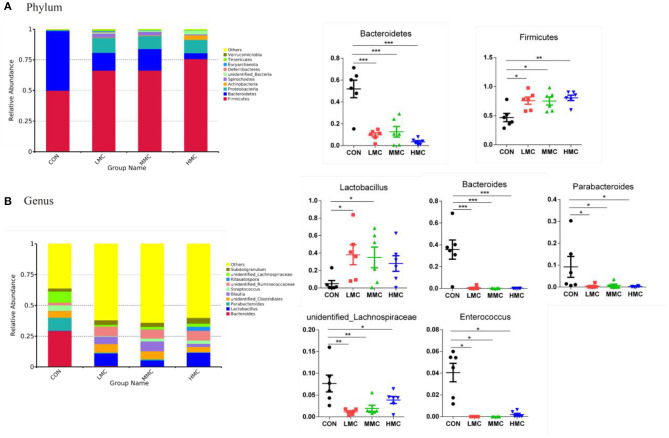
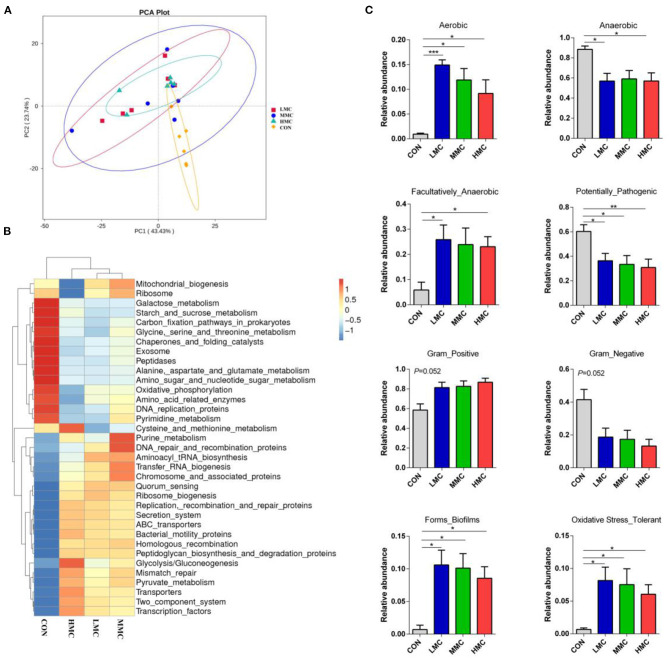
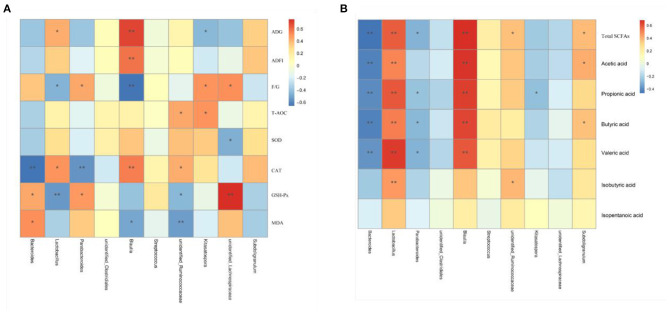
Background: Moutan cortex radicis (MCR), as a common traditional Chinese medicine, has been widely used as an antipyretic, antiseptic, and anti-inflammatory agent in China. Objectives: This study aimed to investigate the effects of dietary MCR supplementation on the antioxidant capacity and intestinal health of the pigs and to explore whether MCR exerts positive effects on intestinal health via regulating nuclear factor kappa-B (NF-κB) signaling pathway and intestinal microbiota. Methods: MCR powder was identified by LC-MS analysis. Selected 32 weaned piglets (21 d of age, 6.37 ± 0.10 kg average BW) were assigned (8 pens/diet, 1 pig/pen) to 4 groups and fed with a corn-soybean basal diet supplemented with 0, 2,000, 4,000, and 8,000 mg/kg MCR for 21 d. After the piglets were sacrificed, antioxidant indices, histomorphology examination, and inflammatory signaling pathway expression were assessed. The 16s RNA sequencing was used to analyze the effects of MCR on the intestinal microbiota structure of piglets. Results: Supplemental 4,000 mg/kg MCR significantly increased (P < 0.05) the average daily weight gain (ADG), average daily feed intake (ADFI), total antioxidative capability, colonic short-chain fatty acids (SCFA) concentrations, and the crypt depth in the jejunum but decreased (P < 0.05) the mRNA expression levels of interferon γ, tumor necrosis factor-α, interleukin-1β, inhibiting kappa-B kinase β (IKKβ), inhibiting nuclear factor kappa-B (IκBα), and NF-κB in the jejunum and ileum. Microbiota sequencing identified that MCR supplementation significantly increased the microbial richness indices (Chao1, ACE, and observed species, P < 0.05) and the relative abundances of Firmicutes and Lactobacillus (P < 0.05), decreased the relative abundances of Bacteroides, Parabacteroides, unidentified_Lachnospiraceae, and Enterococcus (P < 0.05) and had no significant effects on the diversity indices (Shannon and Simpson, P > 0.05). Microbial metabolic phenotypes analysis also showed that the richness of aerobic bacteria and facultative anaerobic bacteria, oxidative stress tolerance, and biofilm forming were significantly increased (P < 0.05), and the richness of anaerobic bacteria and pathogenic potential of gut microbiota were reduced (P < 0.05) by MCR treatment. Regression analysis showed that the optimal MCR supplemental level for growth performance, serum antioxidant capacity, and intestinal health of weaned piglets was 3,420 ~ 4,237 mg/kg. Conclusions: MCR supplementation improved growth performance and serum antioxidant capacity, and alleviated intestinal inflammation by inhibiting the IKKβ/IκBα/NF-κB signaling pathway and affecting intestinal microbiota in weaned piglets.
Keywords: Moutan cortex radicis; NF-κB signaling; antioxidant capacity; intestinal microbiota; weaned piglets.
Copyright © 2021 Bai, Liu, Wang, Shu, Xu, Zhou, Xiong, Huang, Deng, Yin and Liu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Associated data
LinkOut - more resources
Full Text Sources
Miscellaneous