

Lipocalin 2 increases after high-intensity exercise in humans and influences muscle gene expression and differentiation in mice
- PMID: 34224151
- PMCID: PMC9291458
- DOI: 10.1002/jcp.30501
Lipocalin 2 increases after high-intensity exercise in humans and influences muscle gene expression and differentiation in mice
Abstract
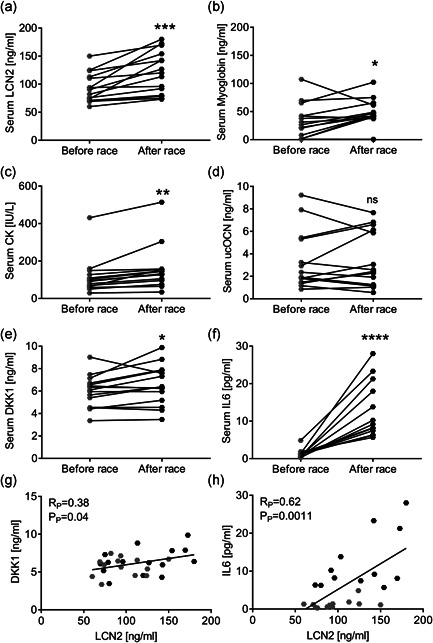
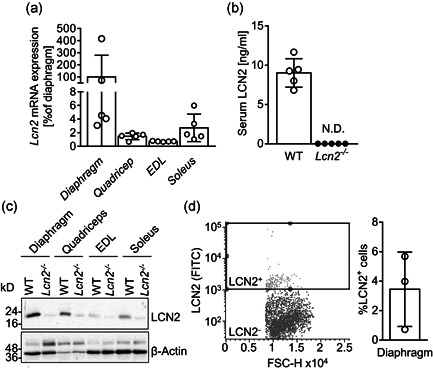
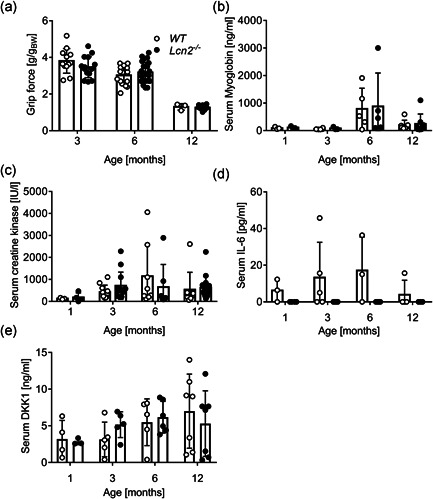
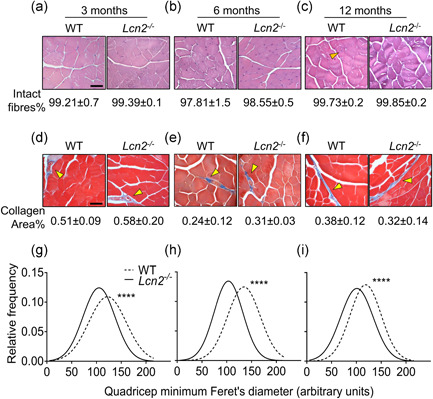
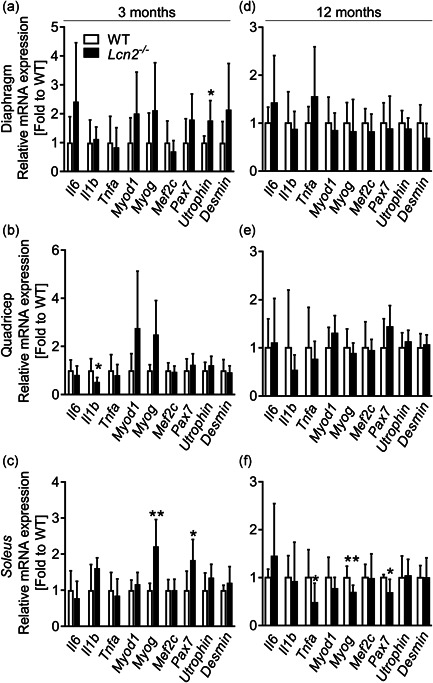
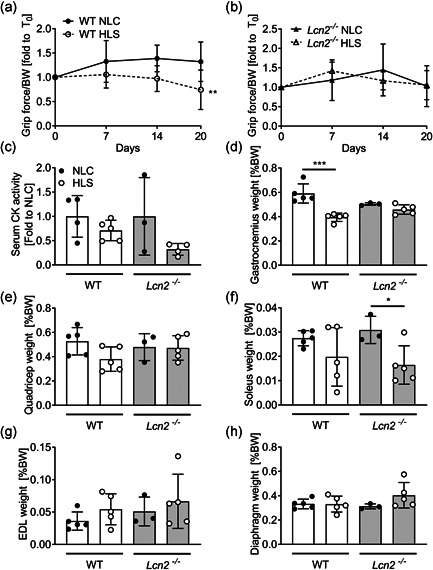
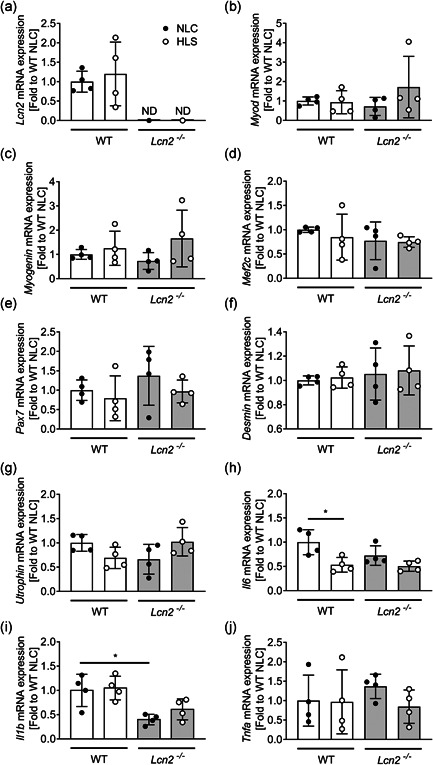
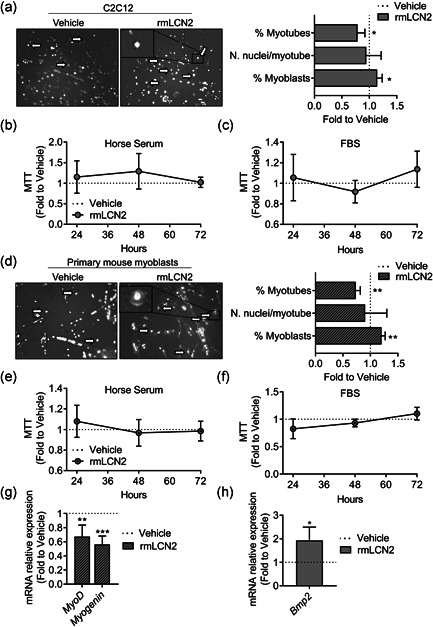
Lipocalin 2 (LCN2) is an adipokine that accomplishes several functions in diverse organs. However, its importance in muscle and physical exercise is currently unknown. We observed that following acute high-intensity exercise ("Gran Sasso d'Italia" vertical run), LCN2 serum levels were increased. The Wnt pathway antagonist, DKK1, was also increased after the run, positively correlating with LCN2, and the same was found for the cytokine Interleukin 6. We, therefore, investigated the involvement of LCN2 in muscle physiology employing an Lcn2 global knockout (Lcn2-/- ) mouse model. Lcn2-/- mice presented with smaller muscle fibres but normal muscle performance (grip strength metre) and muscle weight. At variance with wild type (WT) mice, the inflammatory cytokine Interleukin 6 was undetectable in Lcn2-/- mice at all ages. Intriguingly, Lcn2-/- mice did not lose gastrocnemius and quadriceps muscle mass and muscle performance following hindlimb suspension, while at variance with WT, they lose soleus muscle mass. In vitro, LCN2 treatment reduced the myogenic differentiation of C2C12 and primary mouse myoblasts and influenced their gene expression. Treating myoblasts with LCN2 reduced myogenesis, suggesting that LCN2 may negatively affect muscle physiology when upregulated following high-intensity exercise.
Keywords: adipokines; bone; exercise; lipocalin 2; muscle; myogenesis.
© 2021 The Authors. Journal of Cellular Physiology published by Wiley Periodicals LLC.
Figures
References
-
- Abella, V. , Scotece, M. , Conde, J. , Gómez, R. , Lois, A. , Pino, J. , Gómez‐Reino, J. J. , Lago, F. , Mobasheri, A. , & Gualillo, O. (2015). The potential of lipocalin‐2/NGAL as biomarker for inflammatory and metabolic diseases. Biomarkers, 20(8), 565–571. 10.3109/1354750X.2015.1123354 - DOI - PMC - PubMed
-
- Berger, T. , Togawa, A. , Duncan, G. S. , Elia, A. J. , You‐Ten, A. , Wakeham, A. , Fong, H. E. , Cheung, C. C. , & Mak, T. W. (2006). Lipocalin 2‐deficient mice exhibit increased sensitivity to Escherichia coli infection but not to ischemia‐reperfusion injury. Proceedings of the National Academy of Sciences of the United States of America, 103(6), 1834–1839. 10.1073/pnas.0510847103 - DOI - PMC - PubMed
-
- Capulli, M. , Ponzetti, M. , Maurizi, A. , Gemini‐Piperni, S. , Berger, T. , Mak, T. W. , Teti, A. , & Rucci, N. (2018). A Complex role for Lipocalin 2 in bone metabolism: Global ablation in mice induces osteopenia caused by an altered energy metabolism. Journal of Bone and Mineral Research, 33(6), 1141–1153. 10.1002/jbmr.3406 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
