

Pharmacologic Activation of p53 Triggers Viral Mimicry Response Thereby Abolishing Tumor Immune Evasion and Promoting Antitumor Immunity
- PMID: 34230007
- PMCID: PMC9414294
- DOI: 10.1158/2159-8290.CD-20-1741
Pharmacologic Activation of p53 Triggers Viral Mimicry Response Thereby Abolishing Tumor Immune Evasion and Promoting Antitumor Immunity
Abstract
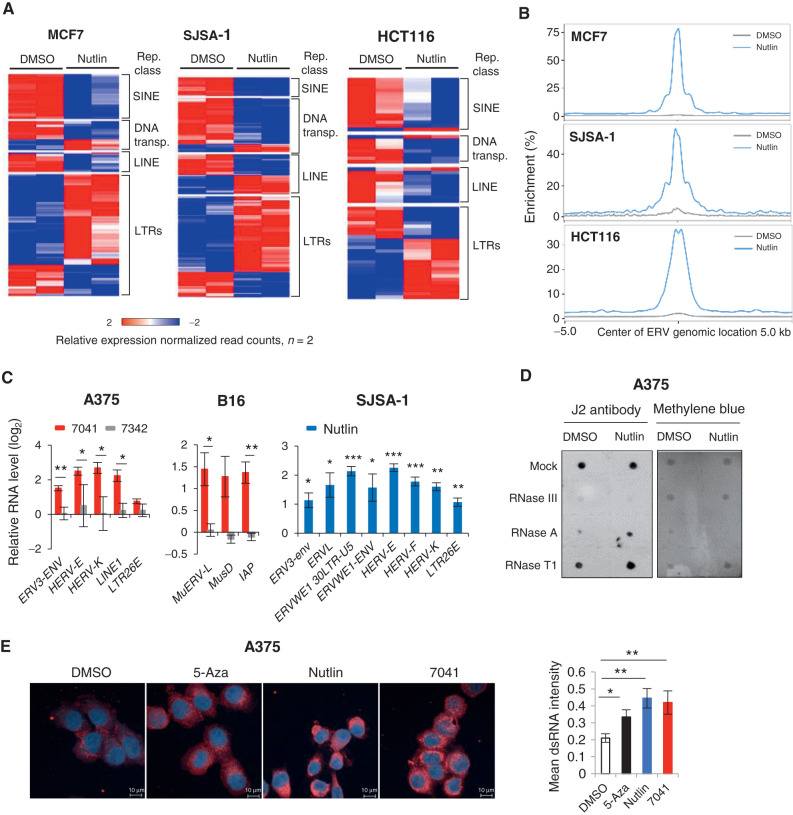
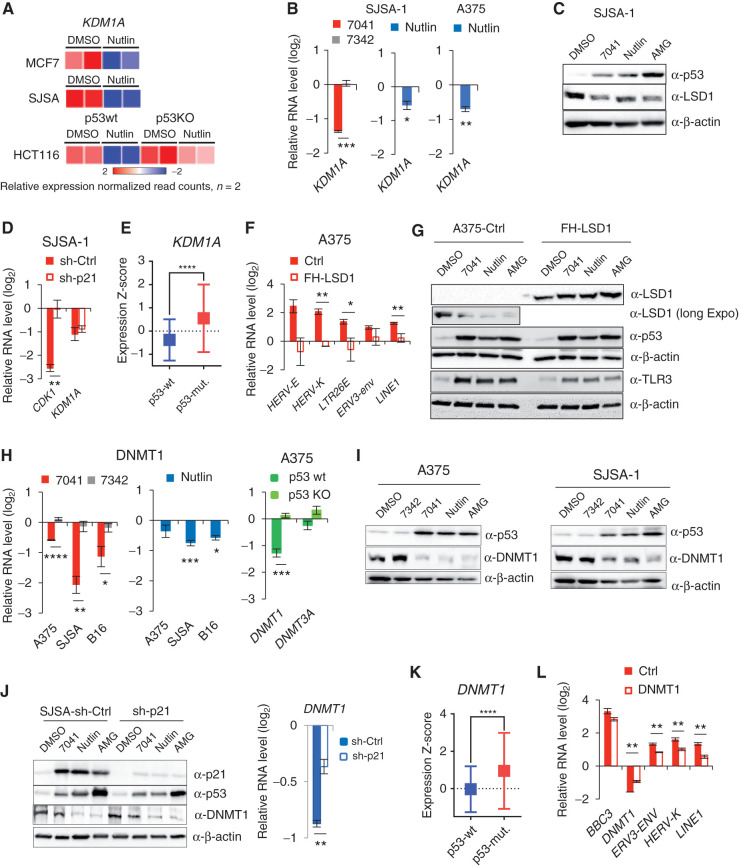
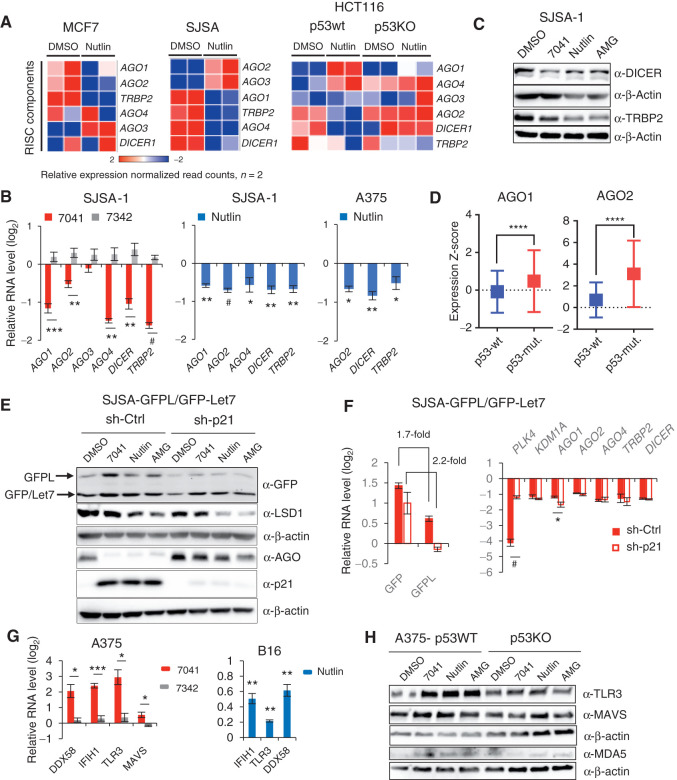
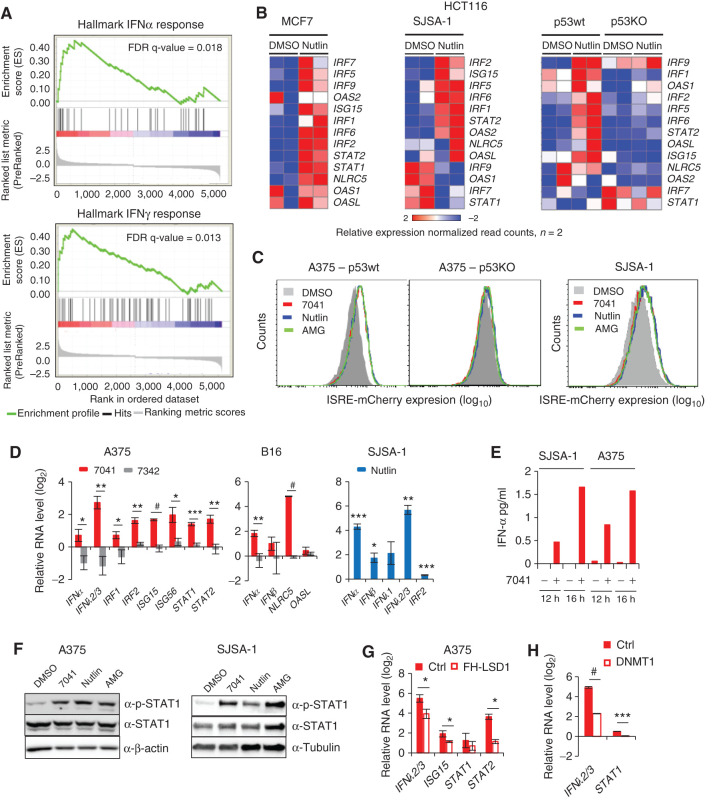
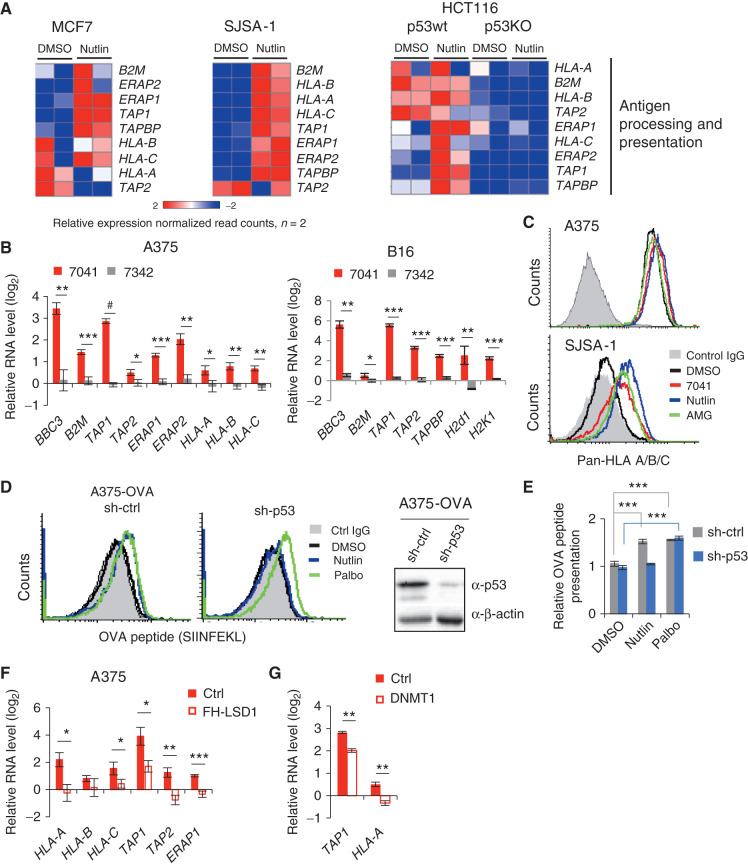
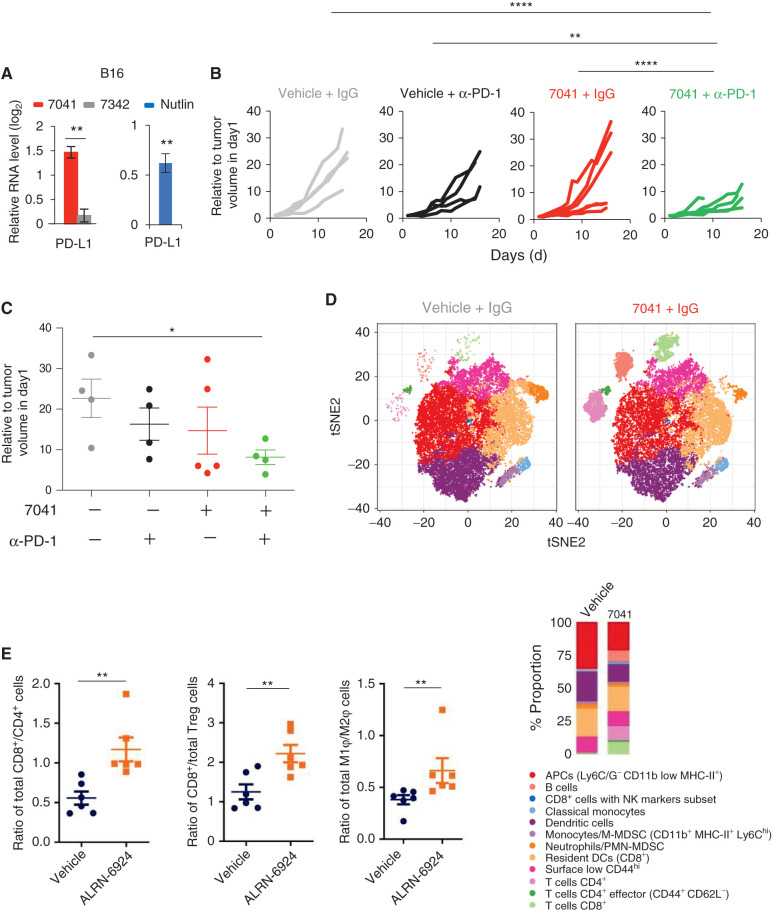
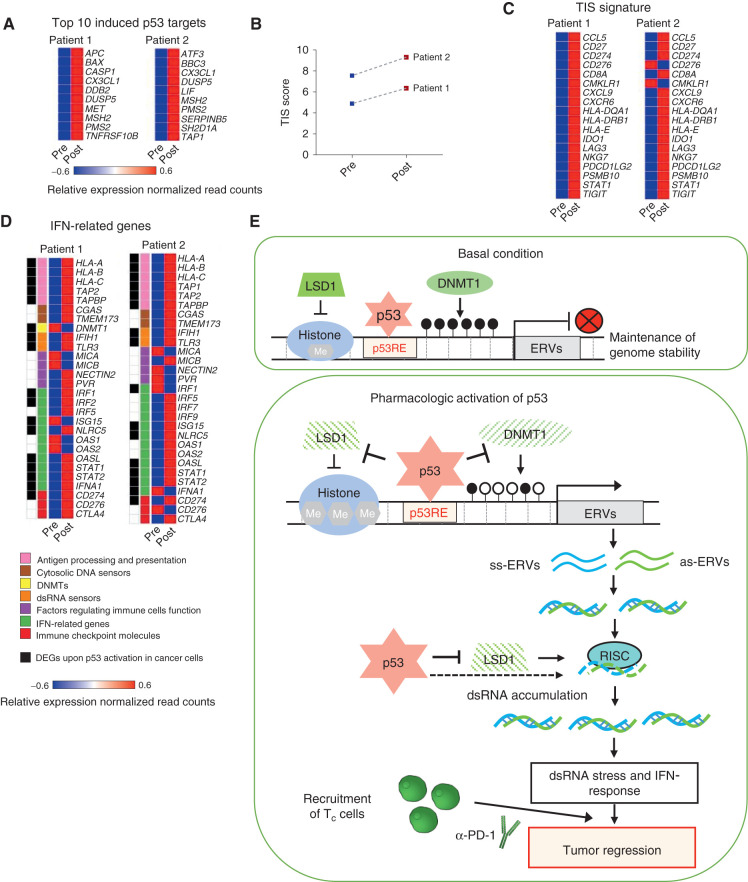
The repression of repetitive elements is an important facet of p53's function as a guardian of the genome. Paradoxically, we found that p53 activated by MDM2 inhibitors induced the expression of endogenous retroviruses (ERV) via increased occupancy on ERV promoters and inhibition of two major ERV repressors, histone demethylase LSD1 and DNA methyltransferase DNMT1. Double-stranded RNA stress caused by ERVs triggered type I/III interferon expression and antigen processing and presentation. Pharmacologic activation of p53 in vivo unleashed the IFN program, promoted T-cell infiltration, and significantly enhanced the efficacy of checkpoint therapy in an allograft tumor model. Furthermore, the MDM2 inhibitor ALRN-6924 induced a viral mimicry pathway and tumor inflammation signature genes in patients with melanoma. Our results identify ERV expression as the central mechanism whereby p53 induction overcomes tumor immune evasion and transforms tumor microenvironment to a favorable phenotype, providing a rationale for the synergy of MDM2 inhibitors and immunotherapy.
Significance: We found that p53 activated by MDM2 inhibitors induced the expression of ERVs, in part via epigenetic factors LSD1 and DNMT1. Induction of IFN response caused by ERV derepression upon p53-targeting therapies provides a possibility to overcome resistance to immune checkpoint blockade and potentially transform "cold" tumors into "hot." This article is highlighted in the In This Issue feature, p. 2945.
Trial registration: ClinicalTrials.gov NCT02264613.
©2021 The Authors; Published by the American Association for Cancer Research.
Figures
Comment in
- doi: 10.1158/2159-8290.CD-11-12-ITI
References
-
- Martins CP, Brown-Swigart L, Evan GI. Modeling the therapeutic efficacy of p53 restoration in tumors. Cell 2006;127:1323–34. - PubMed
-
- Ventura A, Kirsch DG, McLaughlin ME, Tuveson DA, Grimm J, Lintault Let al. . Restoration of p53 function leads to tumour regression in vivo. Nature 2007;445:661–5. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
