

Phase separation in immune signalling
- PMID: 34230650
- PMCID: PMC9674404
- DOI: 10.1038/s41577-021-00572-5
Phase separation in immune signalling
Abstract
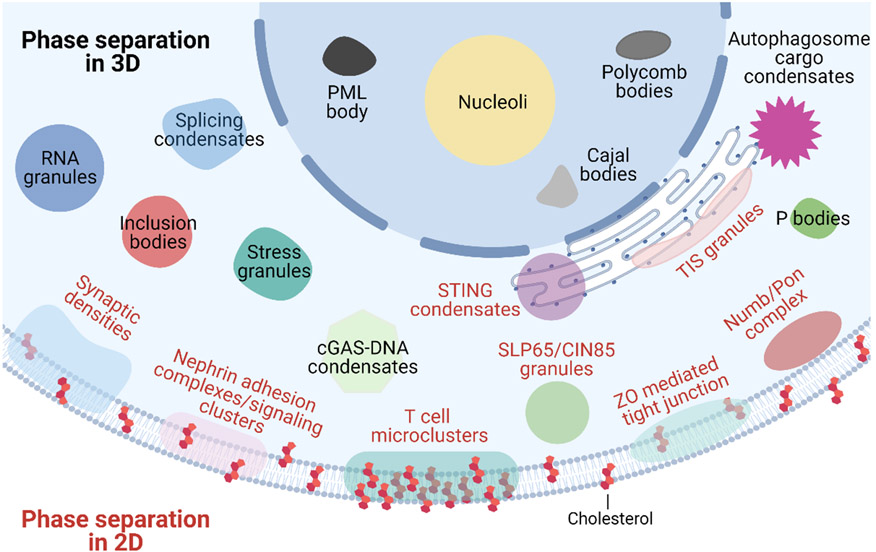
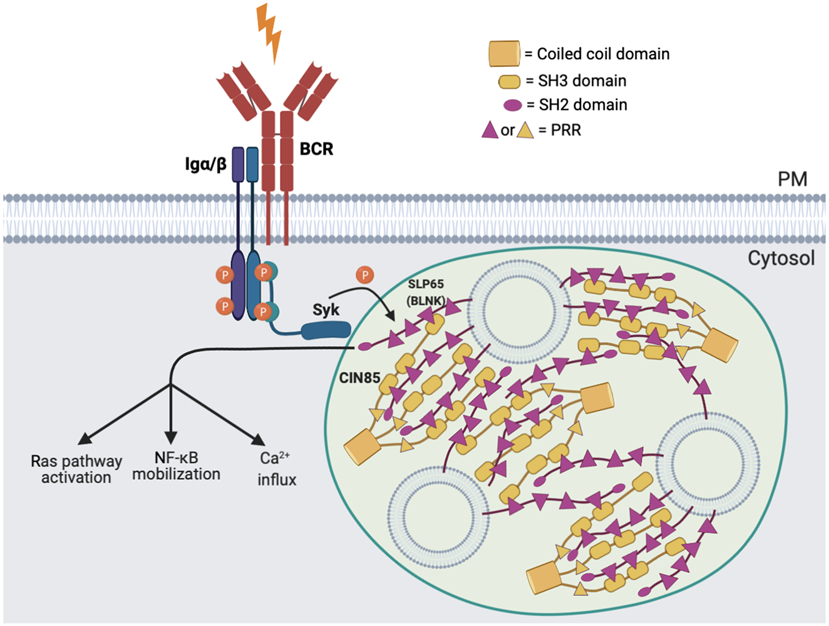
Immune signalling pathways convert pathogenic stimuli into cytosolic events that lead to the resolution of infection. Upon ligand engagement, immune receptors together with their downstream adaptors and effectors undergo substantial conformational changes and spatial reorganization. During this process, nanometre-to-micrometre-sized signalling clusters have been commonly observed that are believed to be hotspots for signal transduction. Because of their large size and heterogeneous composition, it remains a challenge to fully understand the mechanisms by which these signalling clusters form and their functional consequences. Recently, phase separation has emerged as a new biophysical principle for organizing biomolecules into large clusters with fluidic properties. Although the field is still in its infancy, studies of phase separation in immunology are expected to provide new perspectives for understanding immune responses. Here, we present an up-to-date view of how liquid-liquid phase separation drives the formation of signalling condensates and regulates immune signalling pathways, including those downstream of T cell receptor, B cell receptor and the innate immune receptors cyclic GMP-AMP synthase (cGAS)-stimulator of interferon genes (STING) and retinoic acid-inducible gene I protein (RIG-I). We conclude with a summary of the current challenges the field is facing and outstanding questions for future studies.
© 2021. Springer Nature Limited.
Figures


References
-
- Zbinden A, Perez-Berlanga M, De Rossi P & Polymenidou M Phase Separation and Neurodegenerative Diseases: A Disturbance in the Force. Dev Cell 55, 45–68 (2020). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
