

NNT mediates redox-dependent pigmentation via a UVB- and MITF-independent mechanism
- PMID: 34233163
- PMCID: PMC8349839
- DOI: 10.1016/j.cell.2021.06.022
NNT mediates redox-dependent pigmentation via a UVB- and MITF-independent mechanism
Abstract
Ultraviolet (UV) light and incompletely understood genetic and epigenetic variations determine skin color. Here we describe an UV- and microphthalmia-associated transcription factor (MITF)-independent mechanism of skin pigmentation. Targeting the mitochondrial redox-regulating enzyme nicotinamide nucleotide transhydrogenase (NNT) resulted in cellular redox changes that affect tyrosinase degradation. These changes regulate melanosome maturation and, consequently, eumelanin levels and pigmentation. Topical application of small-molecule inhibitors yielded skin darkening in human skin, and mice with decreased NNT function displayed increased pigmentation. Additionally, genetic modification of NNT in zebrafish alters melanocytic pigmentation. Analysis of four diverse human cohorts revealed significant associations of skin color, tanning, and sun protection use with various single-nucleotide polymorphisms within NNT. NNT levels were independent of UVB irradiation and redox modulation. Individuals with postinflammatory hyperpigmentation or lentigines displayed decreased skin NNT levels, suggesting an NNT-driven, redox-dependent pigmentation mechanism that can be targeted with NNT-modifying topical drugs for medical and cosmetic purposes.
Keywords: MITF; UVB; melanosome; nicotinamide nucleotide transhydrogenase; pigmentation; redox regulation.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.E.F. and E.R. have a patent filed on “Methods and compositions for enhancing skin pigmentation” (publication number WO/2016/077817, May 19, 2016.). D.E.F. has a financial interest in Soltego, Inc., a company developing SIK inhibitors for topical skin darkening treatments that might be used for a broad set of human applications. D.E.F.’s interests were reviewed and are managed by Massachusetts General Hospital and Partners HealthCare in accordance with their conflict-of-interest policies. B.P.K. is an inventor on patents and patent applications filed by Mass General Brigham that describe genome engineering technologies. B.P.K. consults for Avectas Inc., ElevateBio, and EcoR1 capital and is an advisor to Acrigen Biosciences. Q.Y.W. is a shareholder in Mymiel Skincare. L.I.Z. is a founder and stockholder of Fate Therapeutics, CAMP4 Therapeutics, Amagma Therapeutics, and Scholar Rock. He is a consultant for Celularity and Cellarity. H.W. is an employee and shareholder of Johnson and Johnson.
Figures
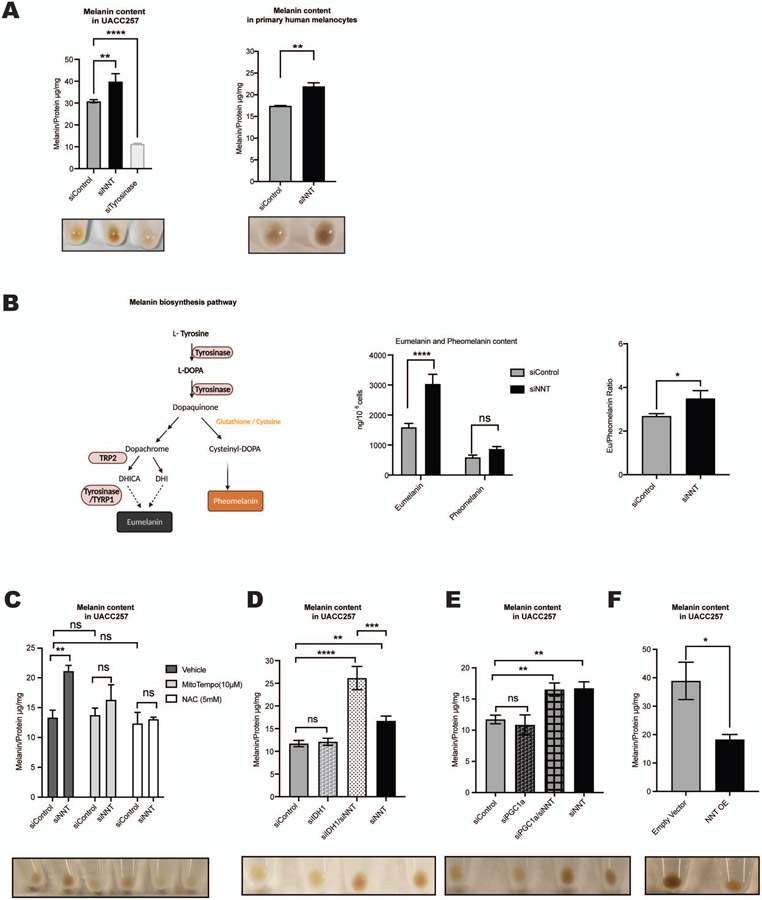
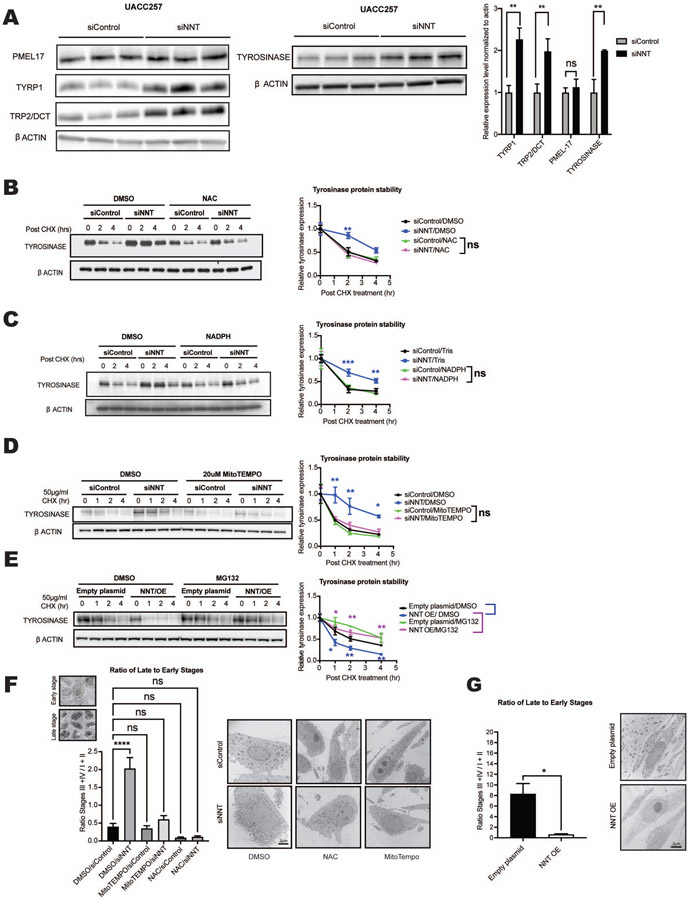
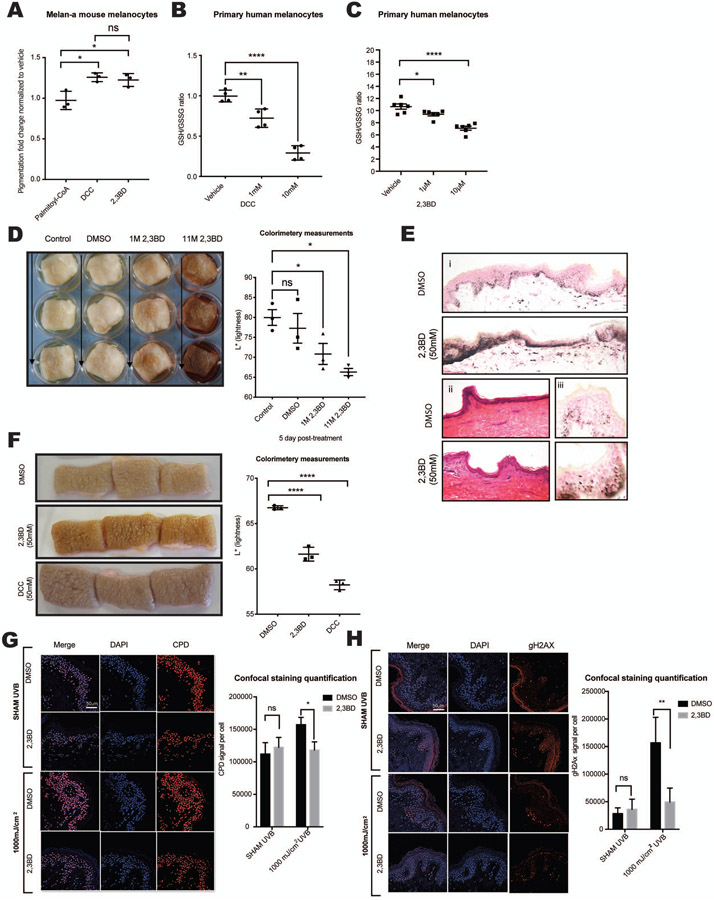
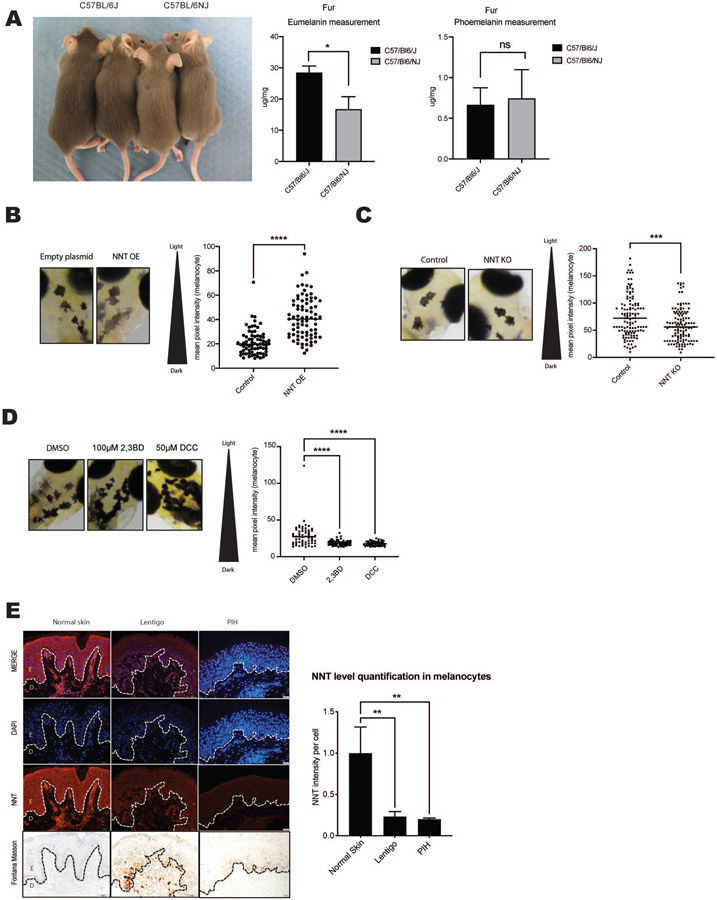
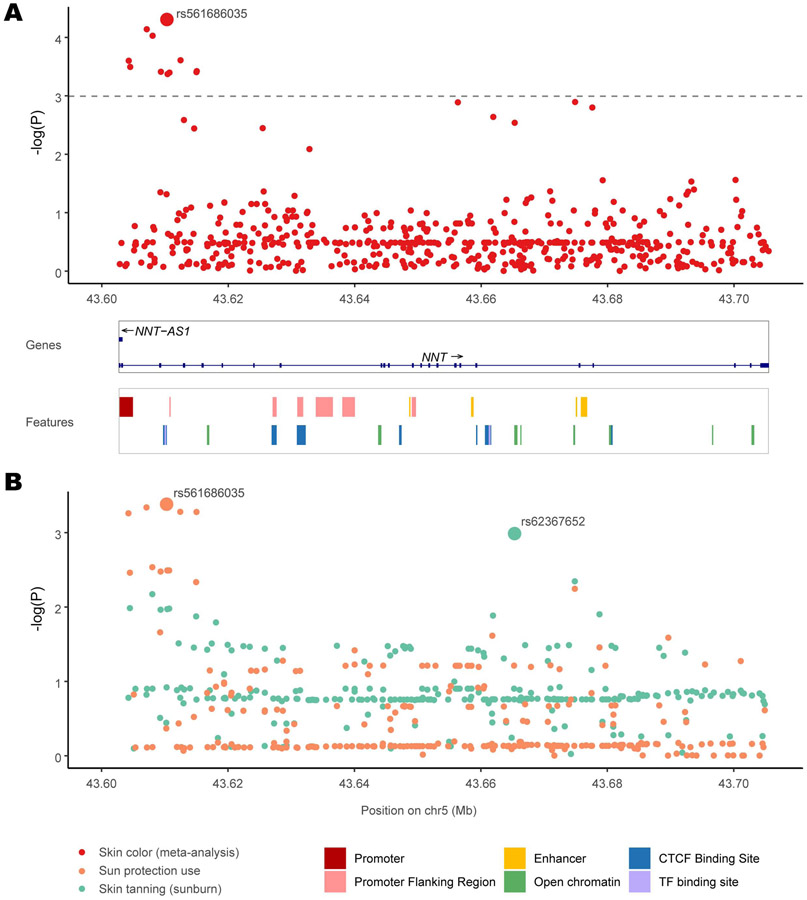
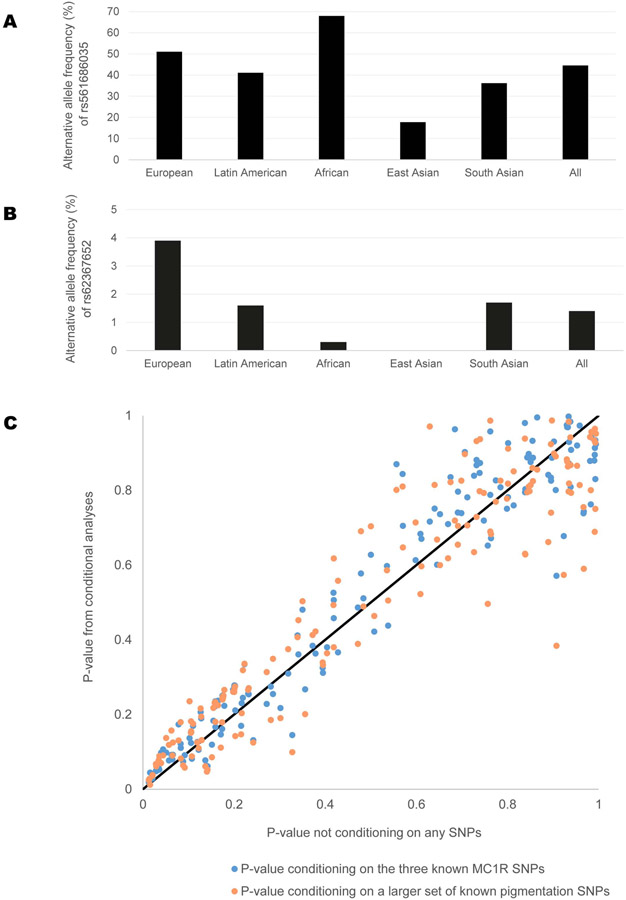
Comment in
-
Commentary on NNT Mediates Redox-Dependent Pigmentation via a UVB-And MITF-Independent Mechanism.J Cell Sci Ther. 2021;12(Suppl 6):316. Epub 2021 Sep 20. J Cell Sci Ther. 2021. PMID: 34956734 Free PMC article. No abstract available.
References
-
- Human Protein Atlas available from http://www.proteinatlas.org.
-
- Adhikari K, Mendoza-Revilla J, Sohail A, Fuentes-Guajardo M, Lampert J, Chacon-Duque JC, Hurtado M, Villegas V, Granja V, Acuna-Alonzo V, et al. (2019). A GWAS in Latin Americans highlights the convergent evolution of lighter skin pigmentation in Eurasia. Nature communications 10, 358. - PMC - PubMed
-
- Allouche J, Bellon N, Saidani M, Stanchina-Chatrousse L, Masson Y, Patwardhan A, Gilles-Marsens F, Delevoye C, Domingues S, Nissan X, et al. (2015). In vitro modeling of hyperpigmentation associated to neurofibromatosis type 1 using melanocytes derived from human embryonic stem cells. Proc Natl Acad Sci U S A 112, 9034–9039. - PMC - PubMed
-
- Arjinpathana N, and Asawanonda P (2012). Glutathione as an oral whitening agent: a randomized, double-blind, placebo-controlled study. J Dermatolog Treat 23, 97–102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AR072304/AR/NIAMS NIH HHS/United States
- R00 CA218870/CA/NCI NIH HHS/United States
- T32 AR007098/AR/NIAMS NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- UL1 TR002541/TR/NCATS NIH HHS/United States
- P01 CA163222/CA/NCI NIH HHS/United States
- R01 AR076241/AR/NIAMS NIH HHS/United States
- R01 CA103846/CA/NCI NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
- R35 GM134957/GM/NIGMS NIH HHS/United States
- BB/I021213/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- P01 HL142494/HL/NHLBI NIH HHS/United States
- P30 ES013508/ES/NIEHS NIH HHS/United States
- R01 CA222871/CA/NCI NIH HHS/United States
- R01 AR043369/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
