

Heparan Sulfate Proteoglycans as Attachment Factor for SARS-CoV-2
- PMID: 34235261
- PMCID: PMC8227597
- DOI: 10.1021/acscentsci.1c00010
Heparan Sulfate Proteoglycans as Attachment Factor for SARS-CoV-2
Abstract
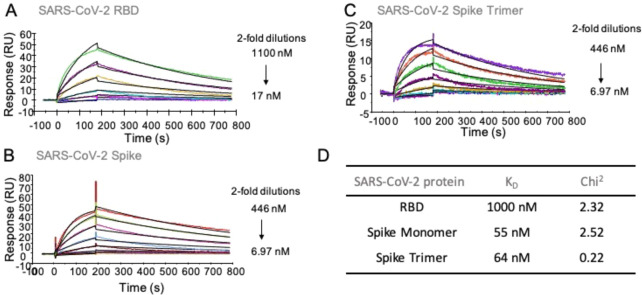
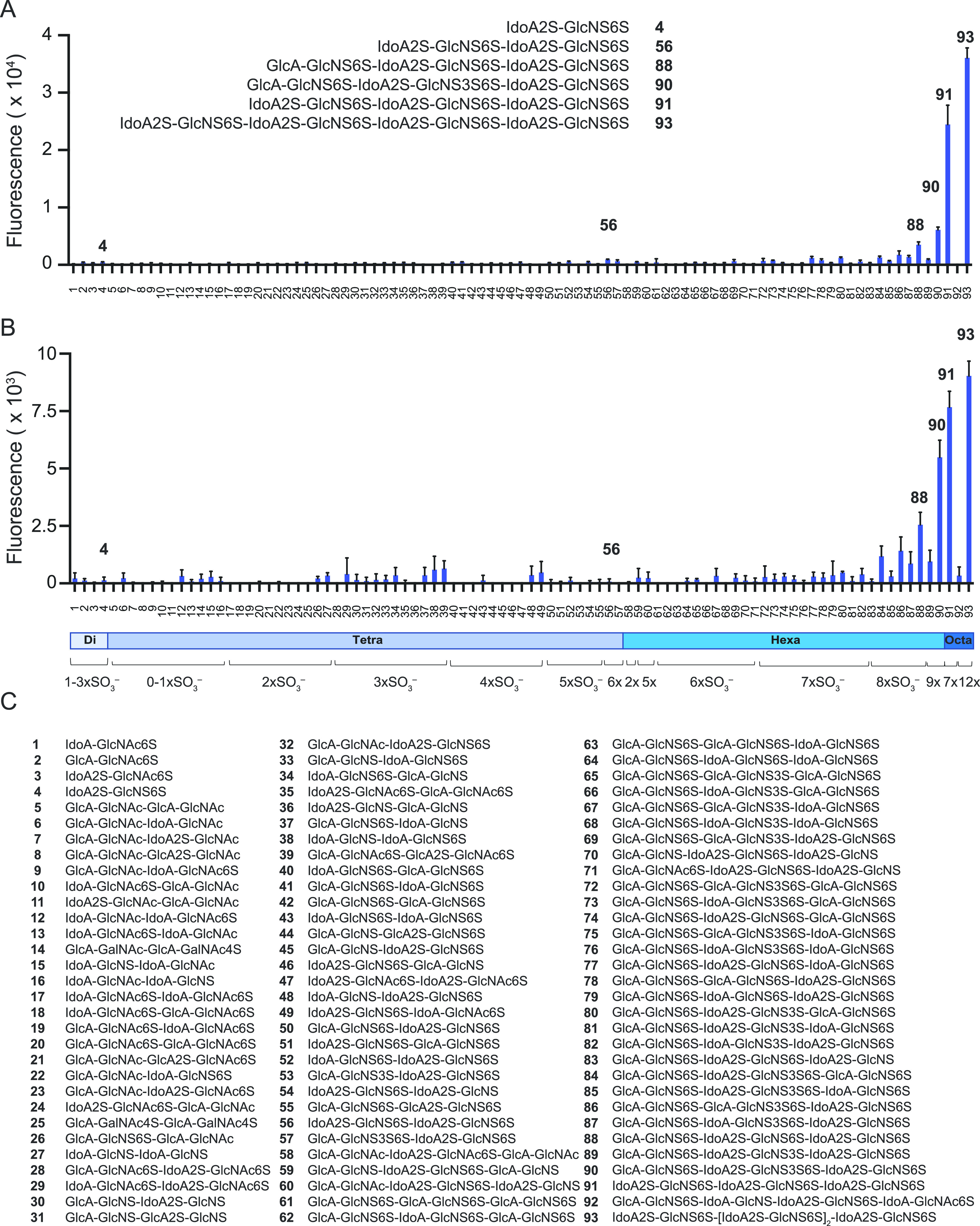
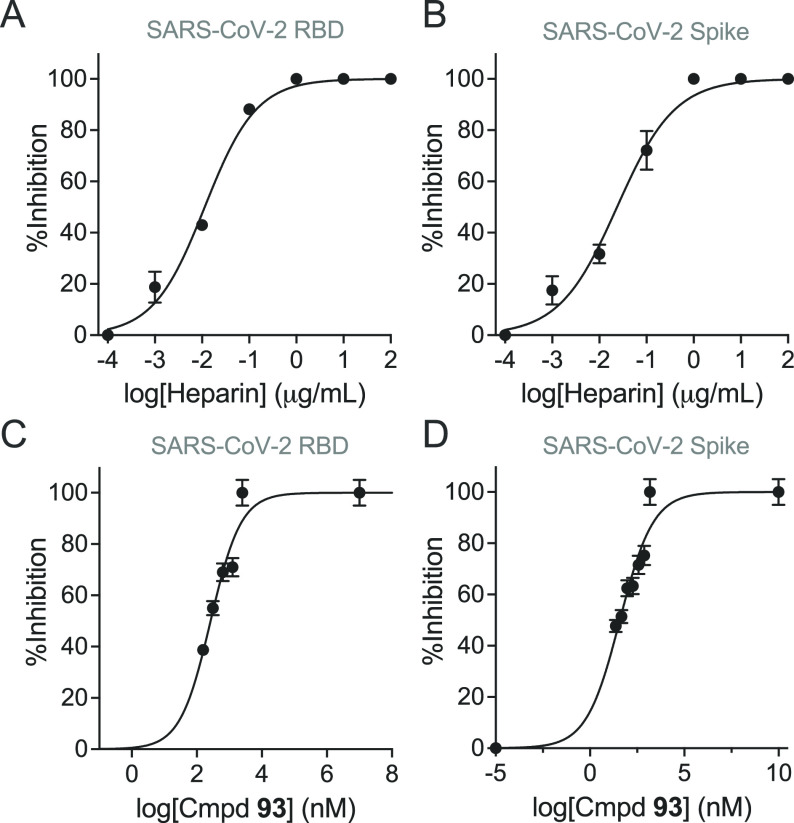
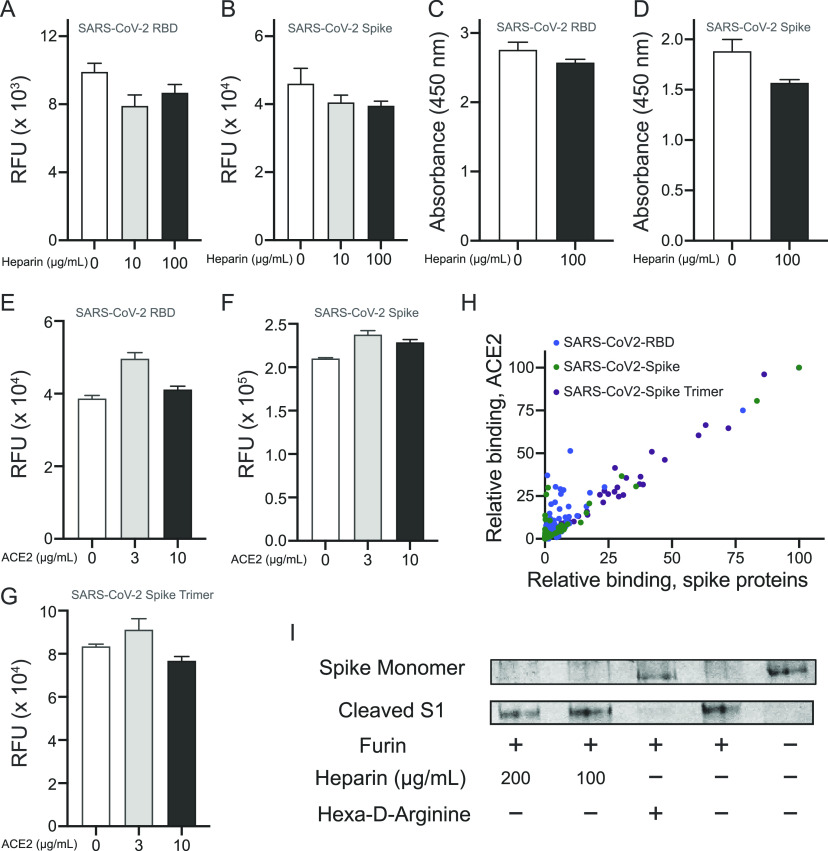
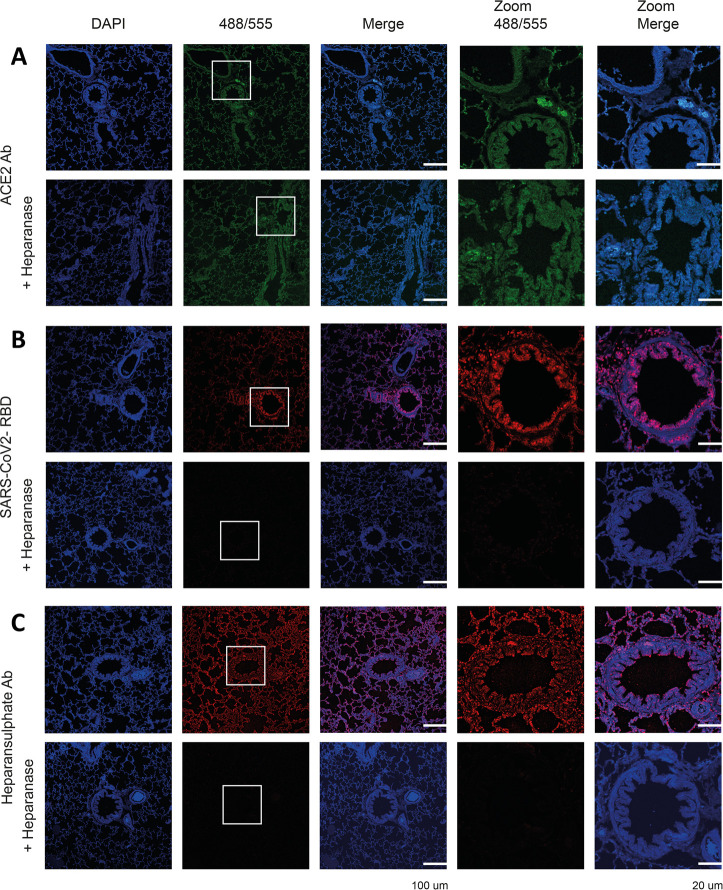
Severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) is causing an unprecedented global pandemic demanding the urgent development of therapeutic strategies. Microarray binding experiments, using an extensive heparan sulfate (HS) oligosaccharide library, showed that the receptor binding domain (RBD) of the spike of SARS-CoV-2 can bind HS in a length- and sequence-dependent manner. A hexasaccharide composed of IdoA2S-GlcNS6S repeating units was identified as the minimal binding epitope. Surface plasmon resonance showed the SARS-CoV-2 spike protein binds with a much higher affinity to heparin (K D = 55 nM) compared to the RBD (K D = 1 μM) alone. It was also found that heparin does not interfere in angiotensin-converting enzyme 2 (ACE2) binding or proteolytic processing of the spike. However, exogenous administered heparin or a highly sulfated HS oligosaccharide inhibited RBD binding to cells. Furthermore, an enzymatic removal of HS proteoglycan from physiological relevant tissue resulted in a loss of RBD binding. The data support a model in which HS functions as the point of initial attachment allowing the virus to travel through the glycocalyx by low-affinity high-avidity interactions to reach the cell membrane, where it can engage with ACE2 for cell entry. Microarray binding experiments showed that ACE2 and HS can simultaneously engage with the RBD, and it is likely no dissociation between HS and RBD is required for binding to ACE2. The results highlight the potential of using HS oligosaccharides as a starting material for therapeutic agent development.
© 2021 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures

Update of
-
Heparan sulfate proteoglycans as attachment factor for SARS-CoV-2.bioRxiv [Preprint]. 2021 Jan 4:2020.05.10.087288. doi: 10.1101/2020.05.10.087288. bioRxiv. 2021. Update in: ACS Cent Sci. 2021 Jun 23;7(6):1009-1018. doi: 10.1021/acscentsci.1c00010. PMID: 32511404 Free PMC article. Updated. Preprint.
References
-
- Monteil V.; Kwon H.; Prado P.; Hagelkruys A.; Wimmer R. A.; Stahl M.; Leopoldi A.; Garreta E.; Hurtado Del Pozo C.; Prosper F.; Romero J. P.; Wirnsberger G.; Zhang H.; Slutsky A. S.; Conder R.; Montserrat N.; Mirazimi A.; Penninger J. M. Inhibition of SARS-CoV-2 infections in engineered human tissues using clinical-grade soluble human ACE2. Cell 2020, 181 (4), 905–913. 10.1016/j.cell.2020.04.004. - DOI - PMC - PubMed
-
- Li W.; Hulswit R. J. G.; Widjaja I.; Raj V. S.; McBride R.; Peng W.; Widagdo W.; Tortorici M. A.; van Dieren B.; Lang Y.; van Lent J. W. M.; Paulson J. C.; de Haan C. A. M.; de Groot R. J.; van Kuppeveld F. J. M.; Haagmans B. L.; Bosch B. J. Identification of sialic acid-binding function for the Middle East respiratory syndrome coronavirus spike glycoprotein. Proc. Natl. Acad. Sci. U. S. A. 2017, 114 (40), E8508–E8517. 10.1073/pnas.1712592114. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
