

Ferritin H deficiency deteriorates cellular iron handling and worsens Salmonella typhimurium infection by triggering hyperinflammation
- PMID: 34236052
- PMCID: PMC8410025
- DOI: 10.1172/jci.insight.141760
Ferritin H deficiency deteriorates cellular iron handling and worsens Salmonella typhimurium infection by triggering hyperinflammation
Abstract
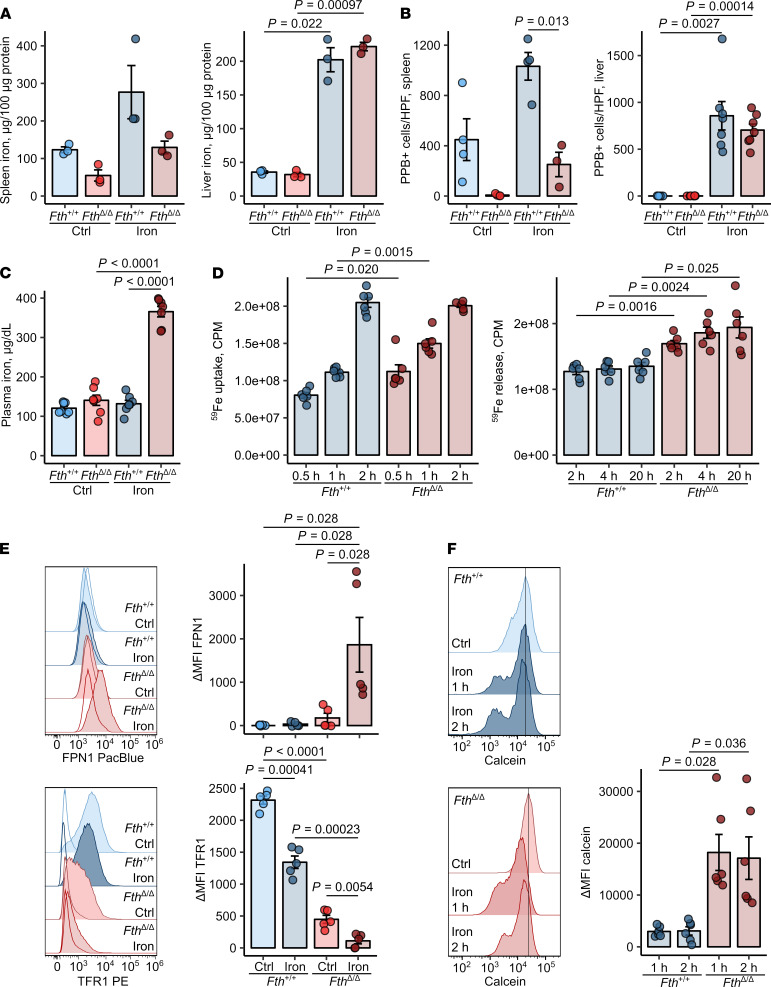
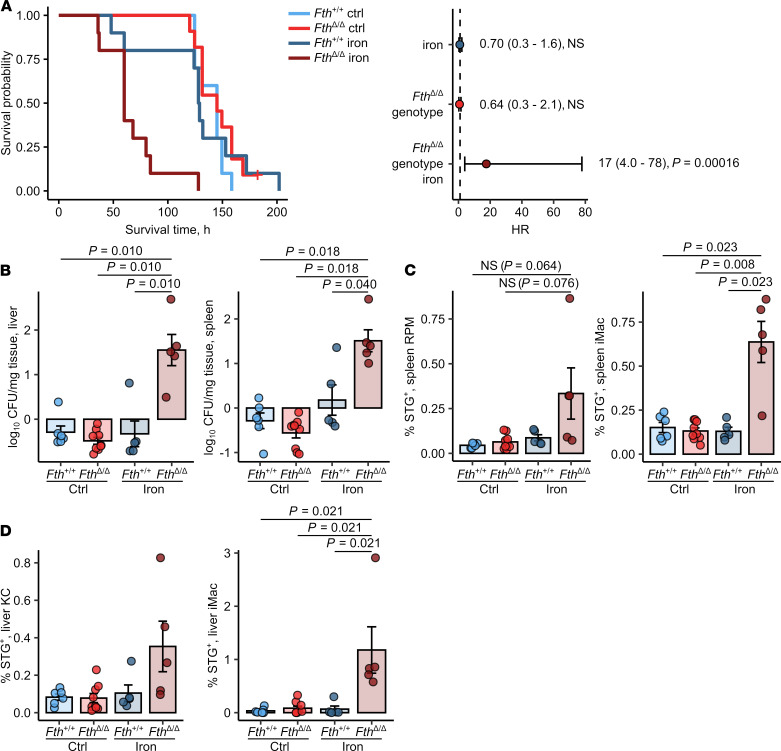
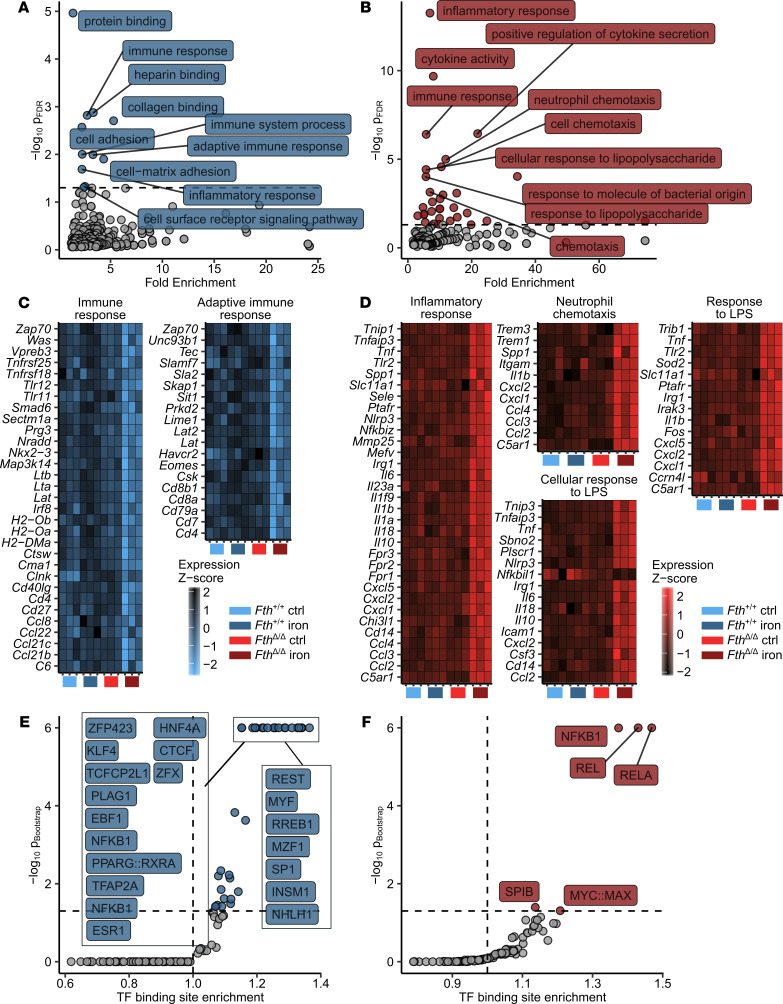
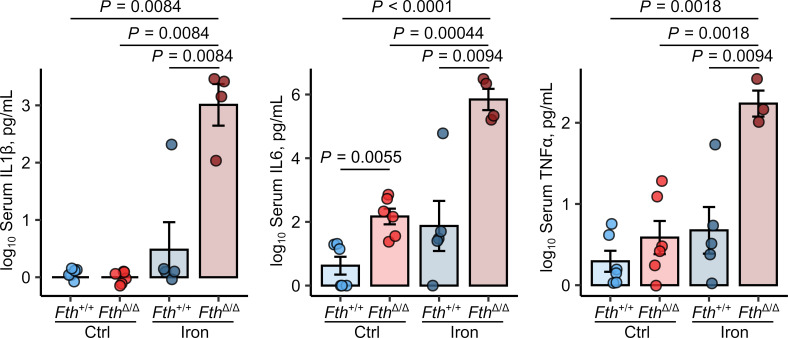
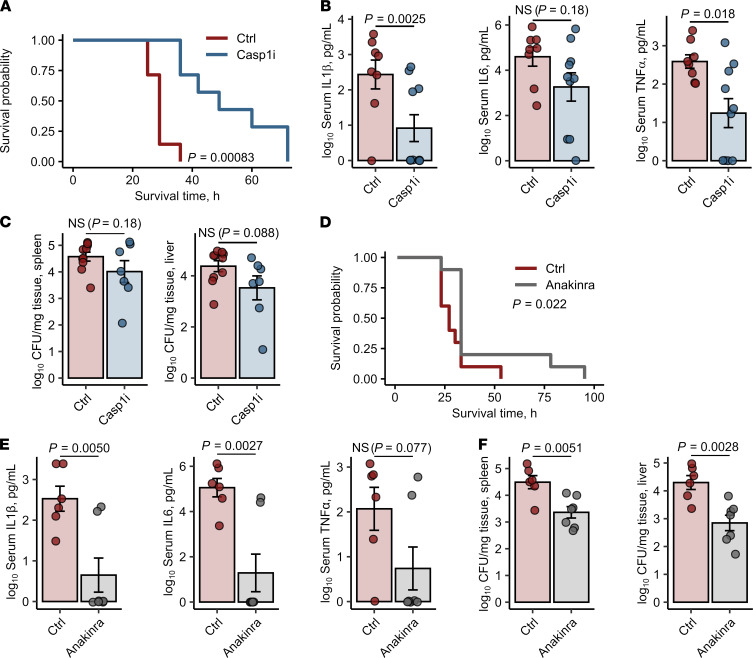
Iron is an essential nutrient for mammals as well as for pathogens. Inflammation-driven changes in systemic and cellular iron homeostasis are central for host-mediated antimicrobial strategies. Here, we studied the role of the iron storage protein ferritin H (FTH) for the control of infections with the intracellular pathogen Salmonella enterica serovar Typhimurium by macrophages. Mice lacking FTH in the myeloid lineage (LysM-Cre+/+Fthfl/fl mice) displayed impaired iron storage capacities in the tissue leukocyte compartment, increased levels of labile iron in macrophages, and an accelerated macrophage-mediated iron turnover. While under steady-state conditions, LysM-Cre+/+Fth+/+ and LysM-Cre+/+Fthfl/fl animals showed comparable susceptibility to Salmonella infection, i.v. iron supplementation drastically shortened survival of LysM-Cre+/+Fthfl/fl mice. Mechanistically, these animals displayed increased bacterial burden, which contributed to uncontrolled triggering of NF-κB and inflammasome signaling and development of cytokine storm and death. Importantly, pharmacologic inhibition of the inflammasome and IL-1β pathways reduced cytokine levels and mortality and partly restored infection control in iron-treated ferritin-deficient mice. These findings uncover incompletely characterized roles of ferritin and cellular iron turnover in myeloid cells in controlling bacterial spread and for modulating NF-κB and inflammasome-mediated cytokine activation, which may be of vital importance in iron-overloaded individuals suffering from severe infections and sepsis.
Keywords: Bacterial infections; Immunology; Infectious disease; Innate immunity.
Conflict of interest statement
Figures
References
-
- Koskenkorva-Frank TS, et al. The complex interplay of iron metabolism, reactive oxygen species, and reactive nitrogen species: insights into the potential of various iron therapies to induce oxidative and nitrosative stress. Free Radic Biol Med. 2013;65:1174–1194. doi: 10.1016/j.freeradbiomed.2013.09.001. - DOI - PubMed
-
- Weinberg ED. Nutritional immunity. Host’s attempt to withold iron from microbial invaders. JAMA. 1975;231(1):39–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
