

Aη-α and Aη-β peptides impair LTP ex vivo within the low nanomolar range and impact neuronal activity in vivo
- PMID: 34238366
- PMCID: PMC8268417
- DOI: 10.1186/s13195-021-00860-1
Aη-α and Aη-β peptides impair LTP ex vivo within the low nanomolar range and impact neuronal activity in vivo
Abstract
Background: Amyloid precursor protein (APP) processing is central to Alzheimer's disease (AD) etiology. As early cognitive alterations in AD are strongly correlated to abnormal information processing due to increasing synaptic impairment, it is crucial to characterize how peptides generated through APP cleavage modulate synapse function. We previously described a novel APP processing pathway producing η-secretase-derived peptides (Aη) and revealed that Aη-α, the longest form of Aη produced by η-secretase and α-secretase cleavage, impaired hippocampal long-term potentiation (LTP) ex vivo and neuronal activity in vivo.
Methods: With the intention of going beyond this initial observation, we performed a comprehensive analysis to further characterize the effects of both Aη-α and the shorter Aη-β peptide on hippocampus function using ex vivo field electrophysiology, in vivo multiphoton calcium imaging, and in vivo electrophysiology.
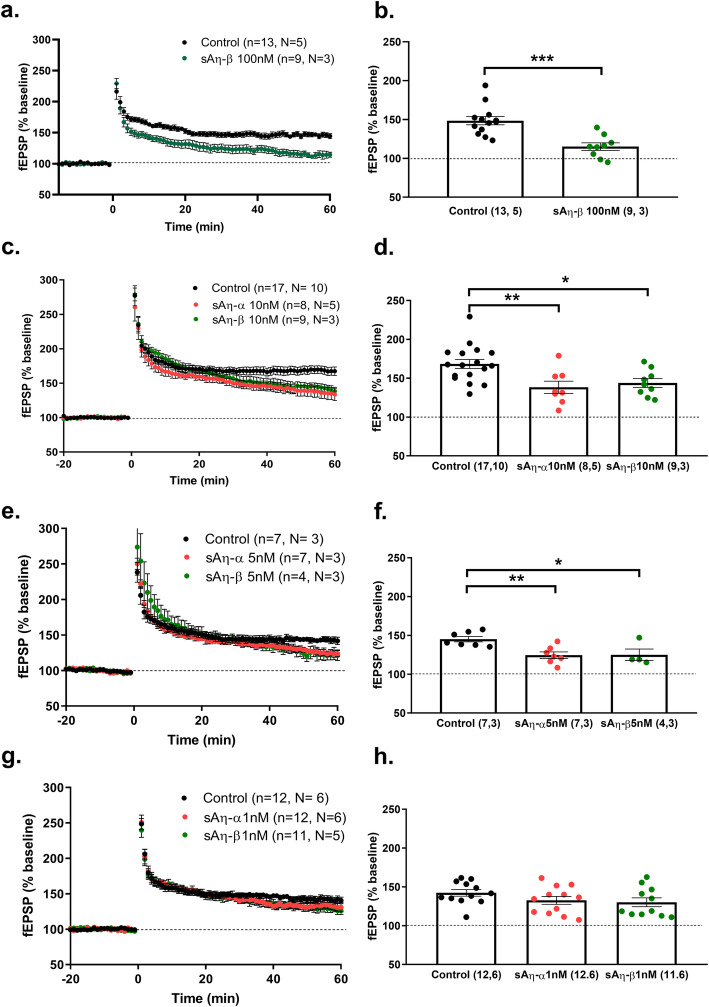
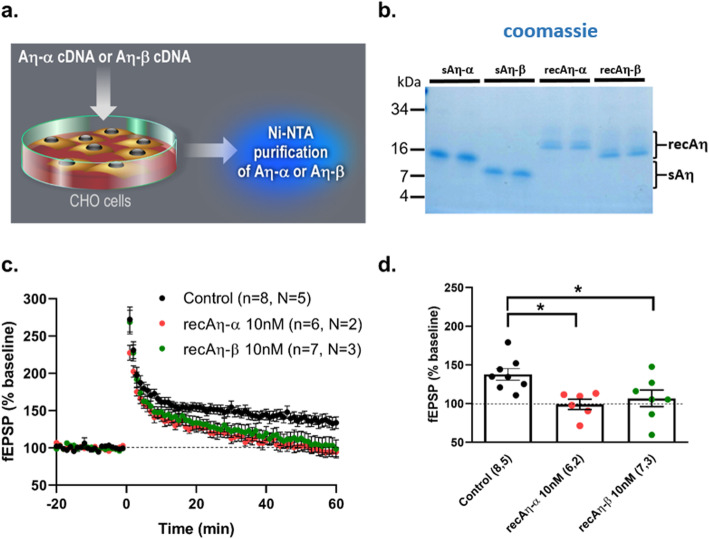
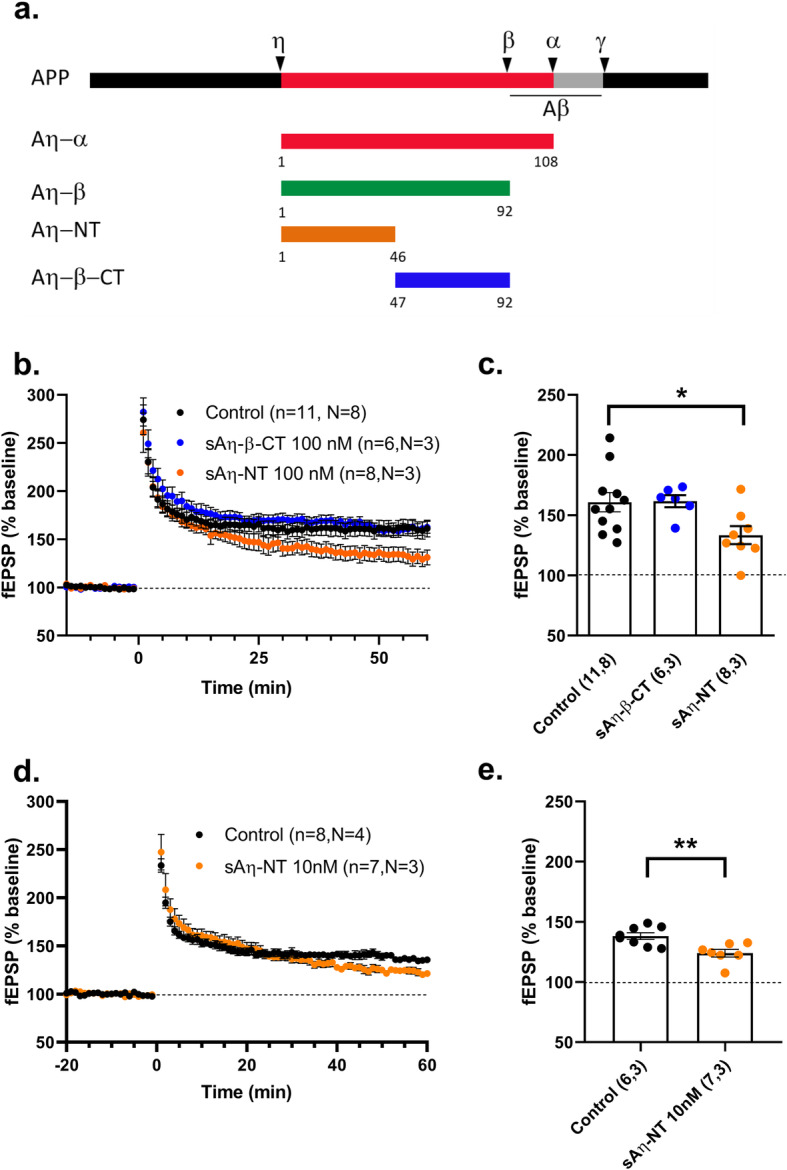
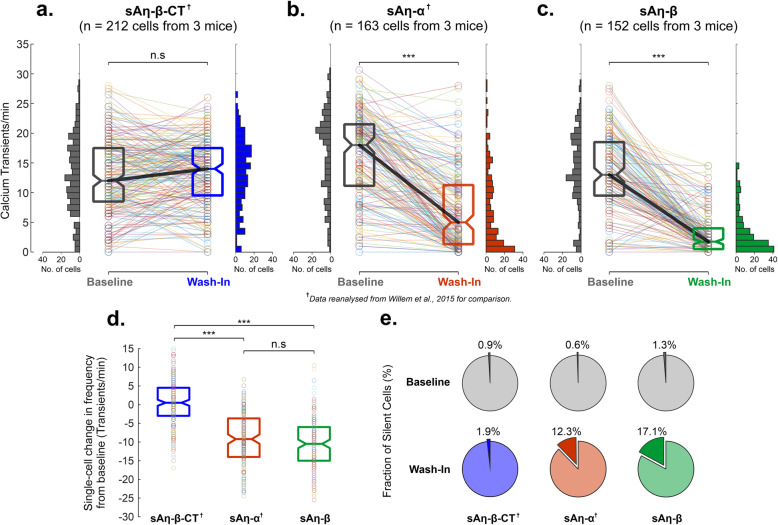
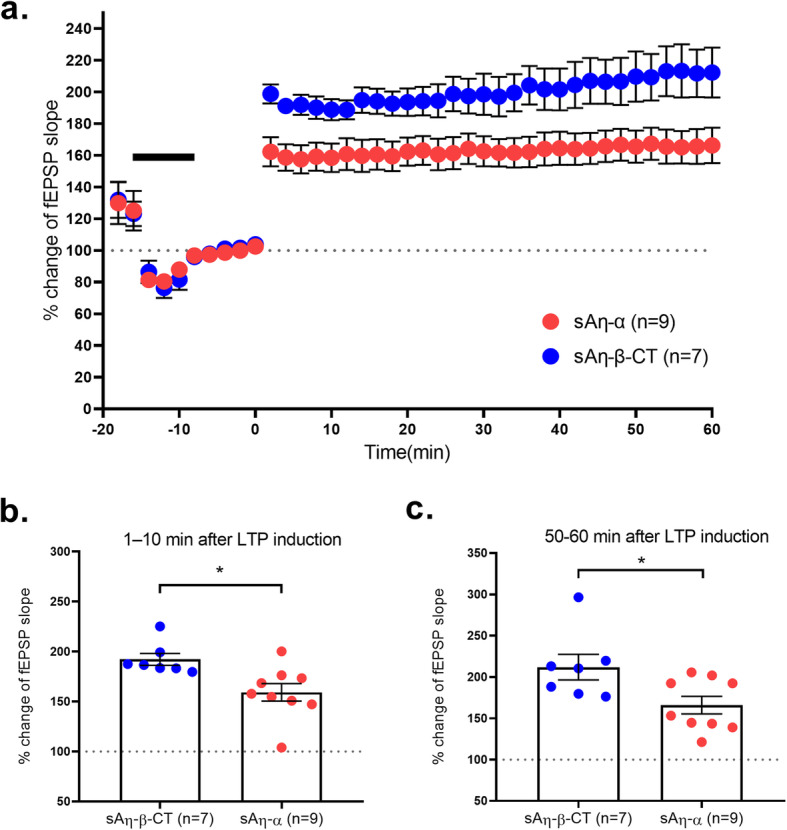
Results: We demonstrate that both synthetic peptides acutely impair LTP at low nanomolar concentrations ex vivo and reveal the N-terminus to be a primary site of activity. We further show that Aη-β, like Aη-α, inhibits neuronal activity in vivo and provide confirmation of LTP impairment by Aη-α in vivo.
Conclusions: These results provide novel insights into the functional role of the recently discovered η-secretase-derived products and suggest that Aη peptides represent important, pathophysiologically relevant, modulators of hippocampal network activity, with profound implications for APP-targeting therapeutic strategies in AD.
Keywords: APP processing; Alzheimer; Electrophysiology; Hippocampus; Synaptic plasticity.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Willem M, Tahirovic S, Busche MA, Ovsepian SV, Chafai M, Kootar S, Hornburg D, Evans LDB, Moore S, Daria A, Hampel H, Müller V, Giudici C, Nuscher B, Wenninger-Weinzierl A, Kremmer E, Heneka MT, Thal DR, Giedraitis V, Lannfelt L, Müller U, Livesey FJ, Meissner F, Herms J, Konnerth A, Marie H, Haass C. eta-Secretase processing of APP inhibits neuronal activity in the hippocampus. Nature. 2015;526(7573):443–447. doi: 10.1038/nature14864. - DOI - PMC - PubMed
-
- Baranger K, Marchalant Y, Bonnet AE, Crouzin N, Carrete A, Paumier JM, Py NA, Bernard A, Bauer C, Charrat E, Moschke K, Seiki M, Vignes M, Lichtenthaler SF, Checler F, Khrestchatisky M, Rivera S. MT5-MMP is a new pro-amyloidogenic proteinase that promotes amyloid pathology and cognitive decline in a transgenic mouse model of Alzheimer’s disease. Cell Mol Life Sci. 2016;73(1):217–236. doi: 10.1007/s00018-015-1992-1. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
