

Genome Analysis Traces Regional Dispersal of Rice in Taiwan and Southeast Asia
- PMID: 34240169
- PMCID: PMC8557449
- DOI: 10.1093/molbev/msab209
Genome Analysis Traces Regional Dispersal of Rice in Taiwan and Southeast Asia
Abstract
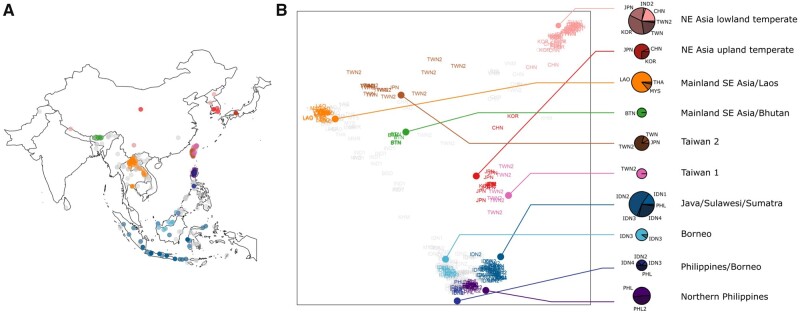
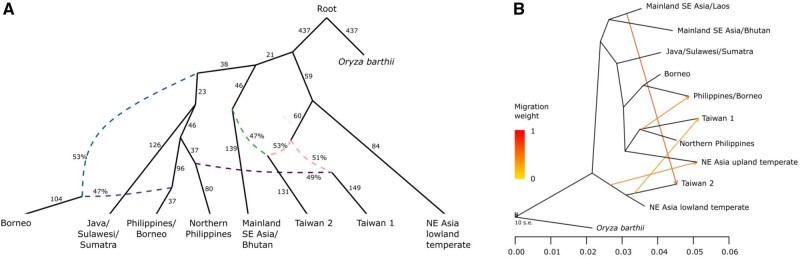
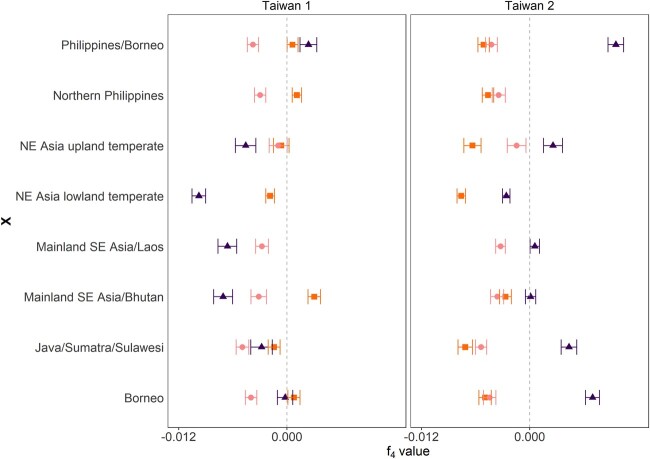
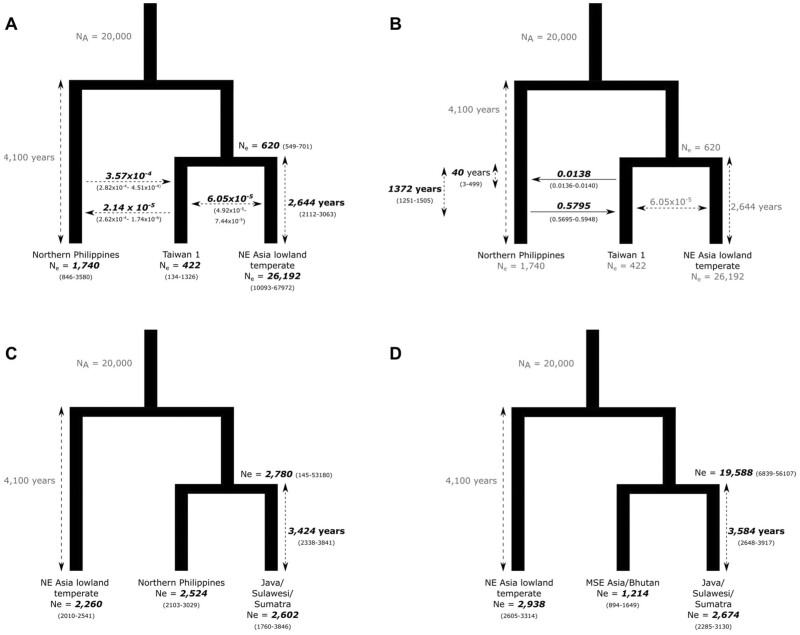
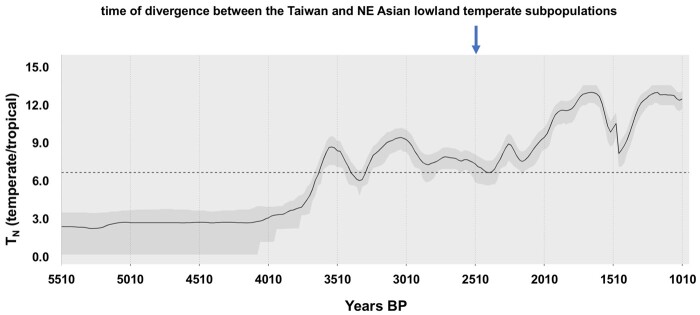
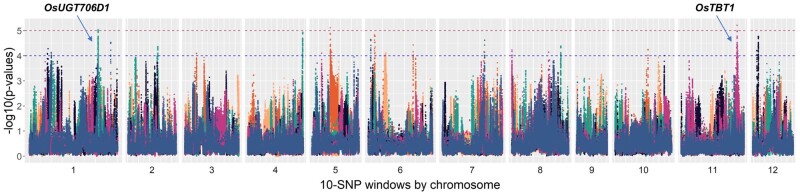
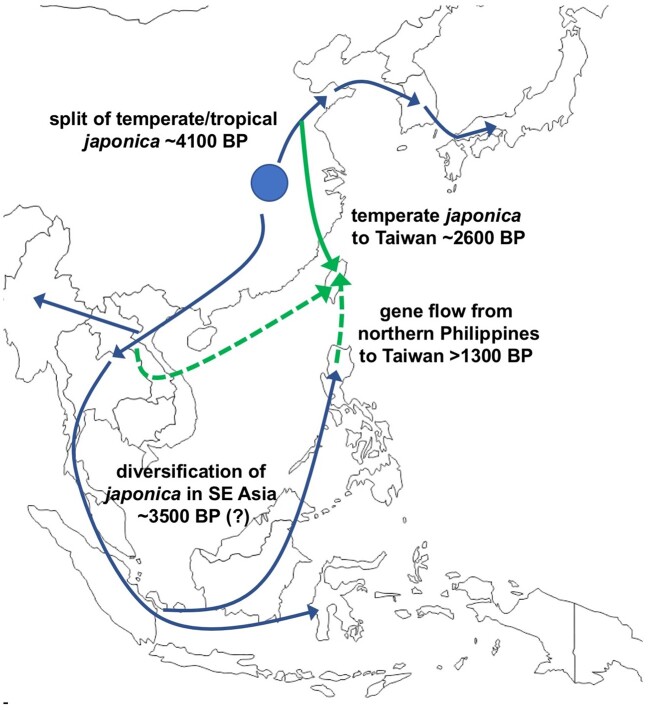
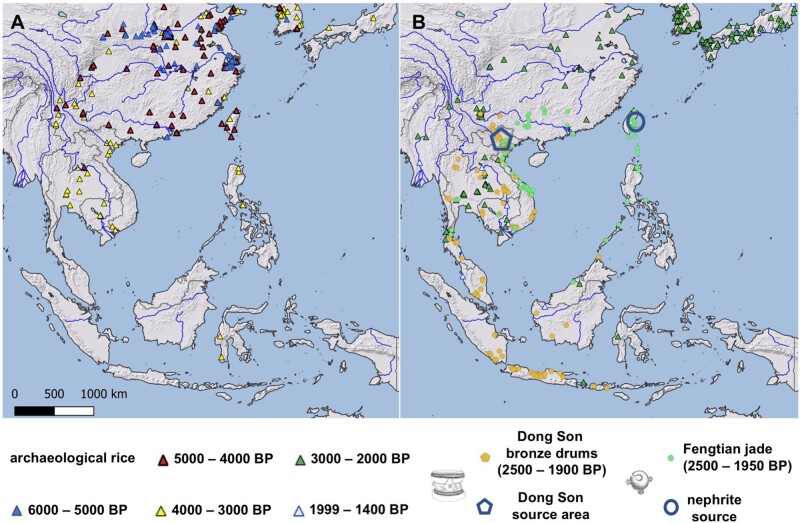
The dispersal of rice (Oryza sativa) following domestication influenced massive social and cultural changes across South, East, and Southeast (SE) Asia. The history of dispersal across islands of SE Asia, and the role of Taiwan and the Austronesian expansion in this process remain largely unresolved. Here, we reconstructed the routes of dispersal of O. sativa ssp. japonica rice to Taiwan and the northern Philippines using whole-genome resequencing of indigenous rice landraces coupled with archaeological and paleoclimate data. Our results indicate that japonica rice found in the northern Philippines diverged from Indonesian landraces as early as 3,500 years before present (BP). In contrast, rice cultivated by the indigenous peoples of the Taiwanese mountains has complex origins. It comprises two distinct populations, each best explained as a result of admixture between temperate japonica that presumably came from northeast Asia, and tropical japonica from the northern Philippines and mainland SE Asia, respectively. We find that the temperate japonica component of these indigenous Taiwan populations diverged from northeast Asia subpopulations at about 2,600 BP, whereas gene flow from the northern Philippines had begun before ∼1,300 BP. This coincides with a period of intensified trade established across the South China Sea. Finally, we find evidence for positive selection acting on distinct genomic regions in different rice subpopulations, indicating local adaptation associated with the spread of japonica rice.
Keywords: Austronesian expansion; admixture; crop evolution; domestication.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Anggraeni ST, Bellwood P, Piper P.. 2014. Neolithic foundations in the Karama valley, West Sulawesi, Indonesia. Antiquity 88:740–756.
-
- Barron A, Datan I, Bellwood P, Wood R, Fuller DQ, Denham T.. 2020. Sherds as archaeobotanical assemblages: Gua Sireh reconsidered. Antiquity 94(377):1325–1336.
-
- Bedford S. 2006. Pieces of the Vanuatu puzzle: archaeology of the north, south and centre. Terra Australis 23. Canberra (ACT; ): Australian National University.
-
- Bellina B. 2017. Khao Sam Kaeo. An early port-city between the Indian Ocean and the South China Sea. Paris: École Française d'Extrême-Orient.
-
- Bellwood P. 1997. Prehistory of the Indo-Malaysian Archipelago. Honolulu (HI: ): University of Hawaii Press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
