

Reduced neutralization of SARS-CoV-2 B.1.617 by vaccine and convalescent serum
- PMID: 34242578
- PMCID: PMC8218332
- DOI: 10.1016/j.cell.2021.06.020
Reduced neutralization of SARS-CoV-2 B.1.617 by vaccine and convalescent serum
Abstract
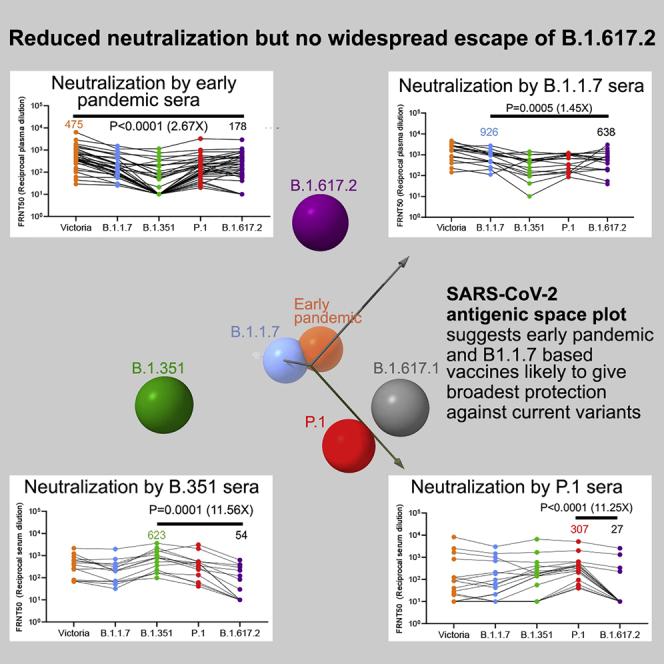
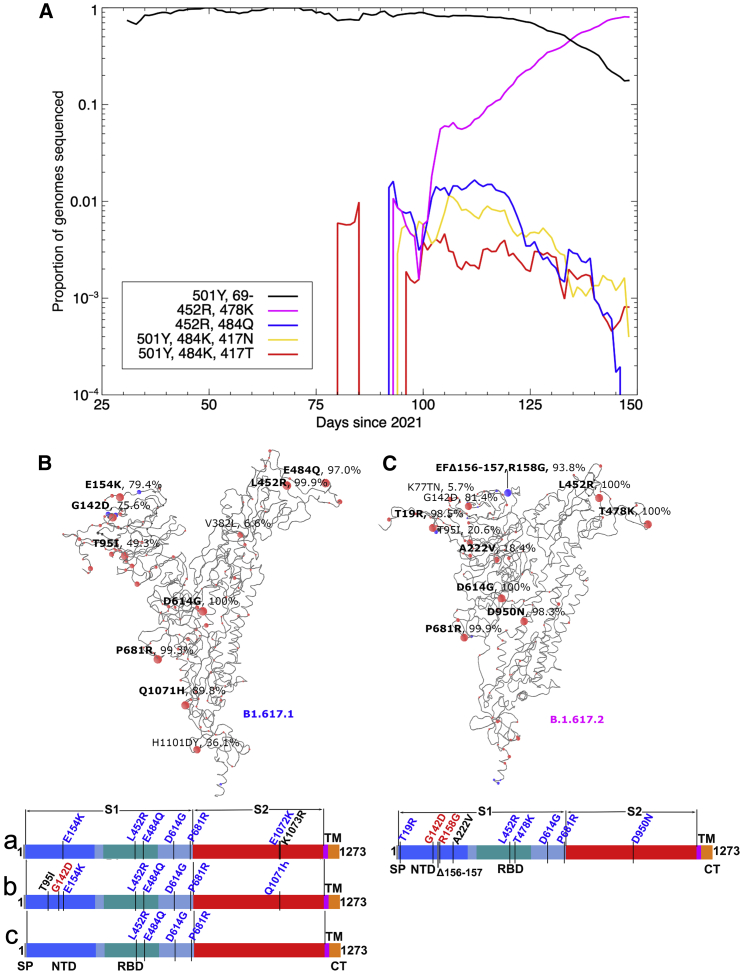
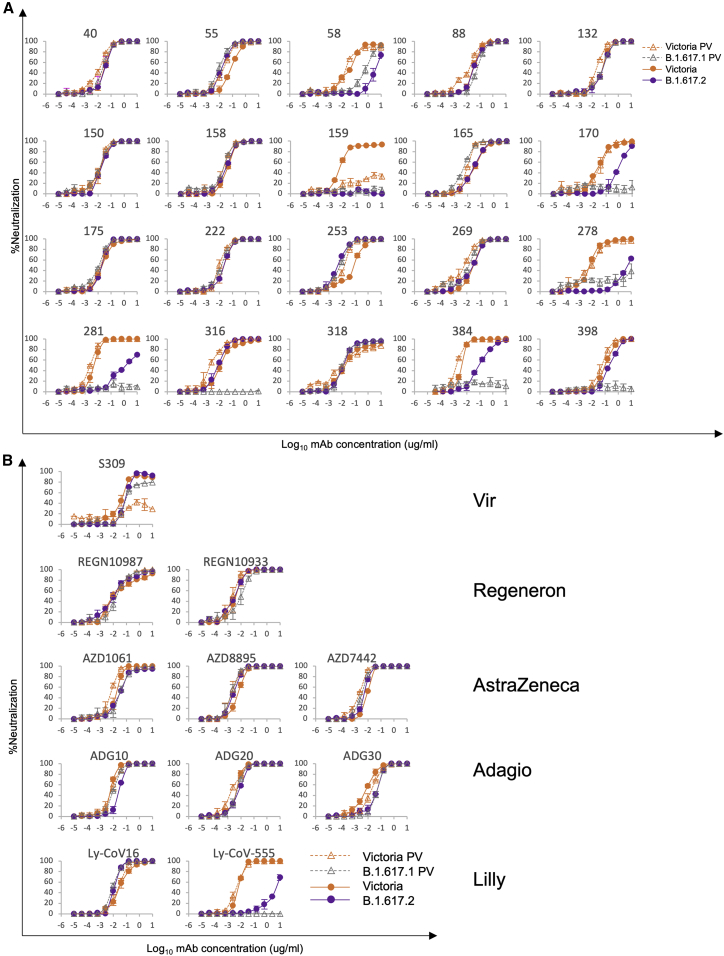
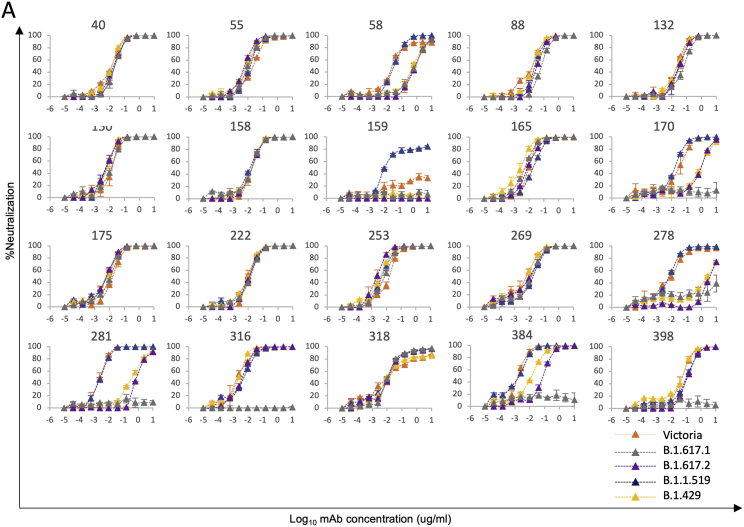
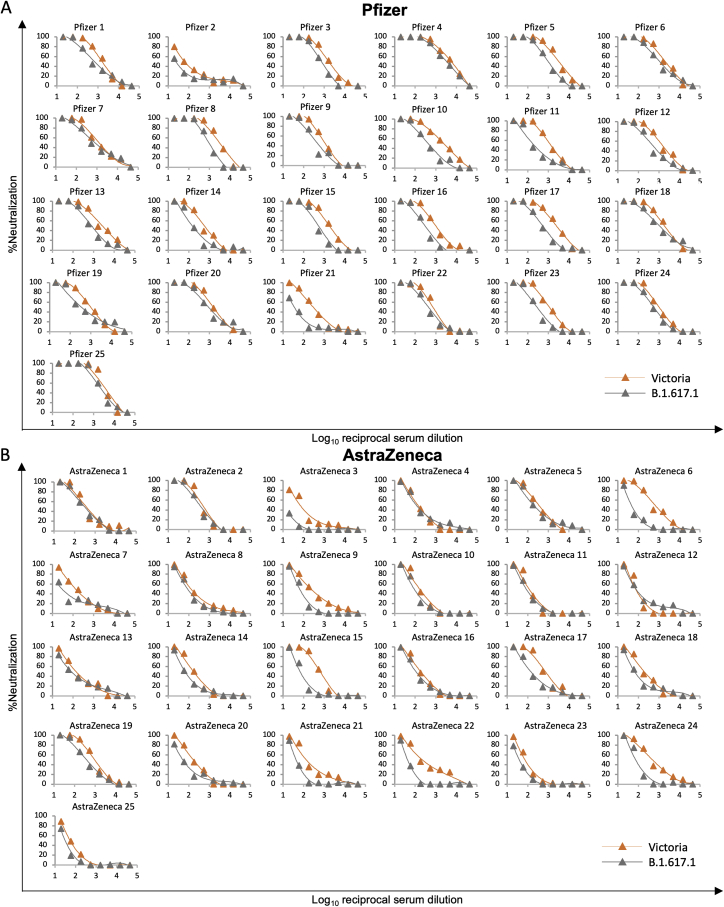
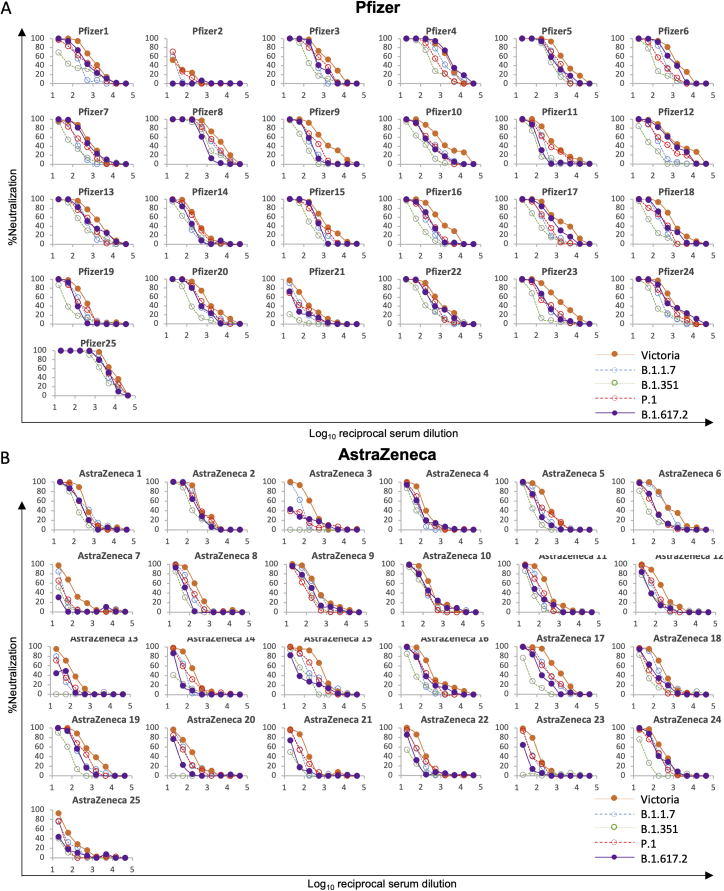
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has undergone progressive change, with variants conferring advantage rapidly becoming dominant lineages, e.g., B.1.617. With apparent increased transmissibility, variant B.1.617.2 has contributed to the current wave of infection ravaging the Indian subcontinent and has been designated a variant of concern in the United Kingdom. Here we study the ability of monoclonal antibodies and convalescent and vaccine sera to neutralize B.1.617.1 and B.1.617.2, complement this with structural analyses of Fab/receptor binding domain (RBD) complexes, and map the antigenic space of current variants. Neutralization of both viruses is reduced compared with ancestral Wuhan-related strains, but there is no evidence of widespread antibody escape as seen with B.1.351. However, B.1.351 and P.1 sera showed markedly more reduction in neutralization of B.1.617.2, suggesting that individuals infected previously by these variants may be more susceptible to reinfection by B.1.617.2. This observation provides important new insights for immunization policy with future variant vaccines in non-immune populations.
Keywords: B.1.617; Delta variant; Receptor-binding-domain; SARS-CoV-2; antibody; escape; neutralization; structure; vaccine; variant.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.R.S. is on the GSK Vaccines Scientific Advisory Board. Oxford University holds intellectual property related to the Oxford-AstraZeneca vaccine. A.J.P. is Chair of UK Department Health and Social Care’s (DHSC) Joint Committee on Vaccination & Immunisation (JCVI) but does not participate in the JCVI COVID19 committee and is a member of the WHO’s SAGE. The views expressed in this article do not necessarily represent the views of DHSC, JCVI, or WHO. The University of Oxford has entered into a partnership with AstraZeneca on coronavirus vaccine development. The University of Oxford has protected intellectual property disclosed in this publication. S.C.G. is co-founder of Vaccitech (collaborators in the early development of this vaccine candidate) and is named as an inventor on a patent covering use of ChAdOx1-vectored vaccines and a patent application covering this SARS-CoV-2 vaccine (PCT/GB2012/000467). T.L. is named as an inventor on a patent application covering this SARS-CoV-2 vaccine and was a consultant to Vaccitech for an unrelated project during the conduct of the study.
Figures
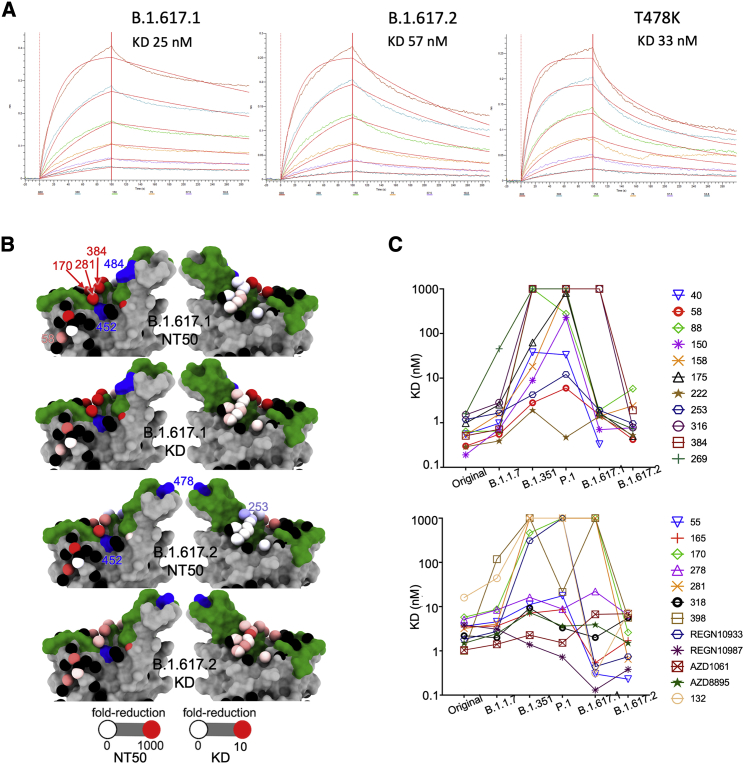
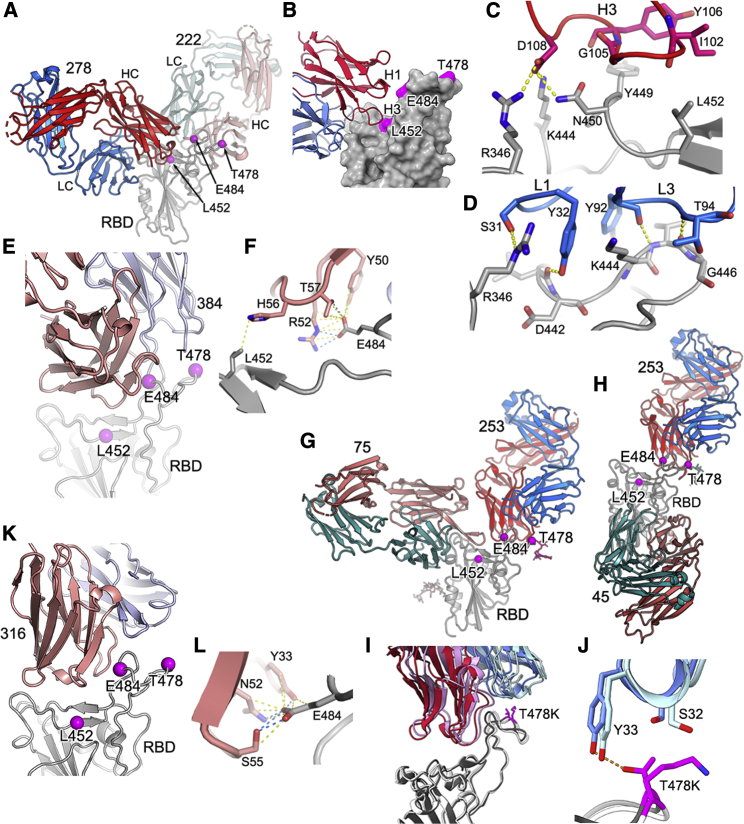
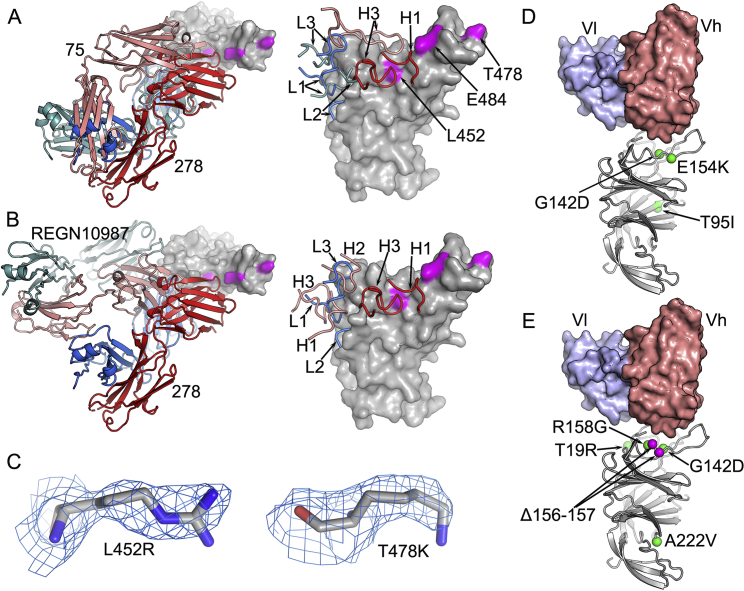
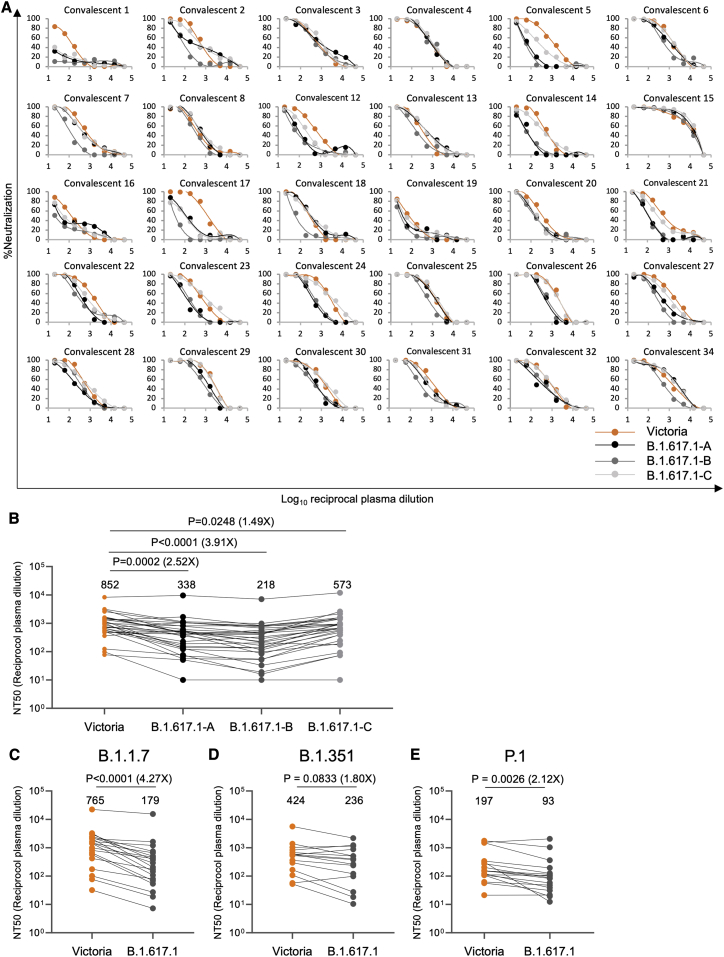
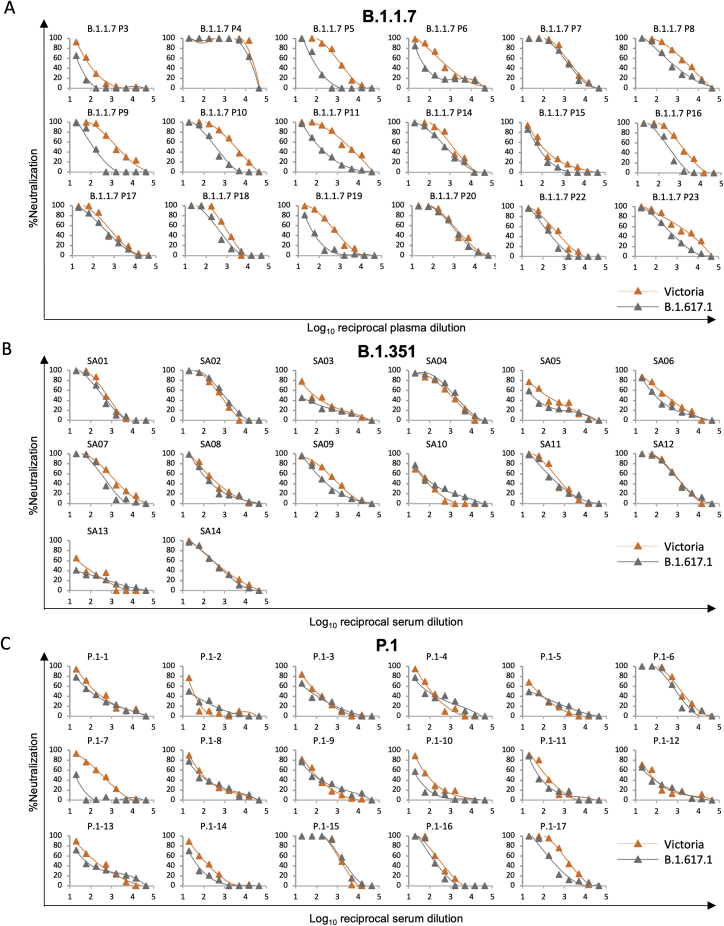
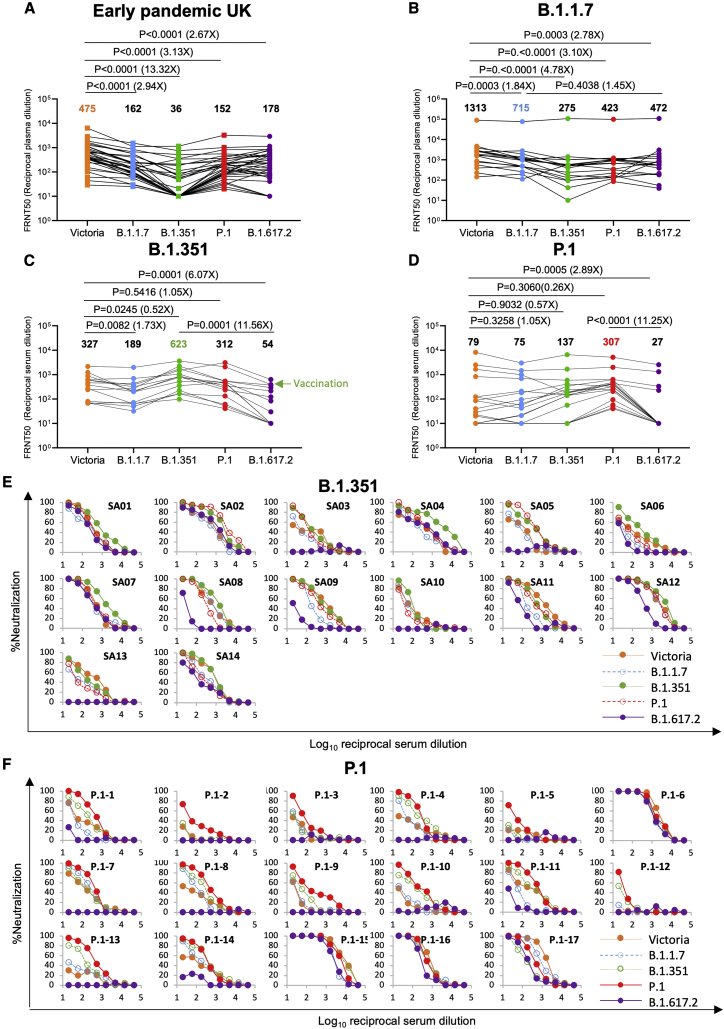
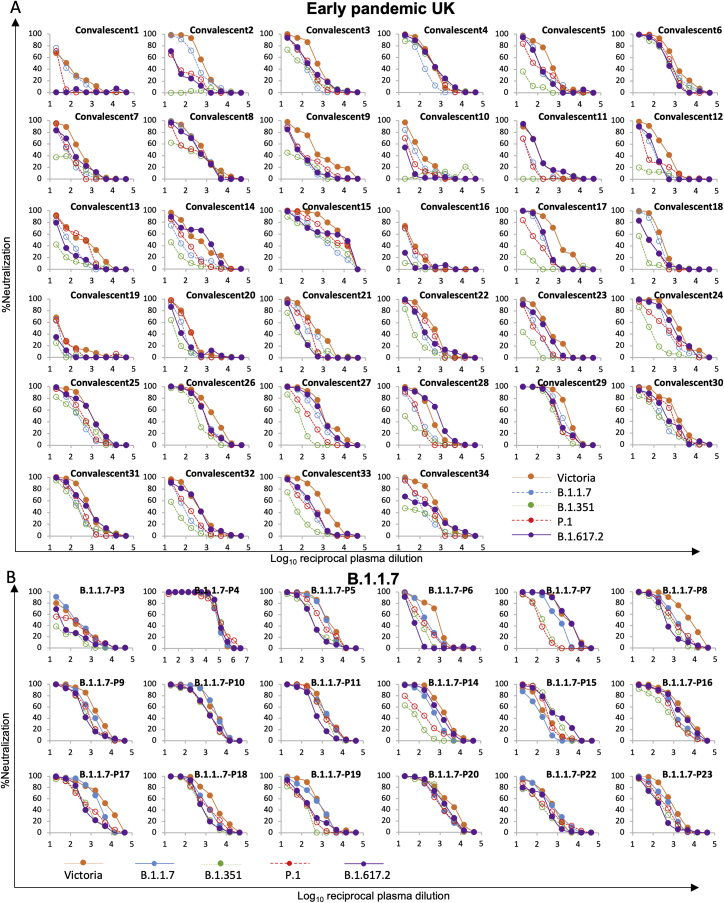
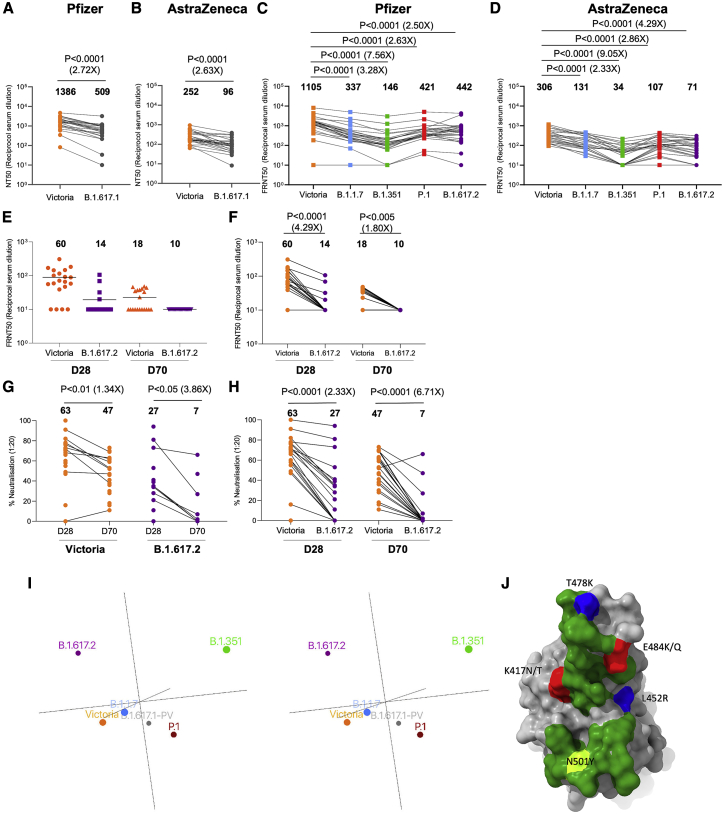
References
Publication types
MeSH terms
Substances
Grants and funding
- 203224/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- MC_PC_19055/MRC_/Medical Research Council/United Kingdom
- MR/V001329/1/MRC_/Medical Research Council/United Kingdom
- MR/N00065X/1/MRC_/Medical Research Council/United Kingdom
- MC_UU_00008/11/MRC_/Medical Research Council/United Kingdom
- MC_PC_20016/MRC_/Medical Research Council/United Kingdom
- MR/V028448/1/MRC_/Medical Research Council/United Kingdom
- MR/L018942/1/MRC_/Medical Research Council/United Kingdom
- MC_PC_19060/MRC_/Medical Research Council/United Kingdom
- G0600520/MRC_/Medical Research Council/United Kingdom
- 109965/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- G1001046/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
