The force loading rate drives cell mechanosensing through both reinforcement and cytoskeletal softening
- PMID: 34244477
- PMCID: PMC8270983
- DOI: 10.1038/s41467-021-24383-3
The force loading rate drives cell mechanosensing through both reinforcement and cytoskeletal softening
Abstract
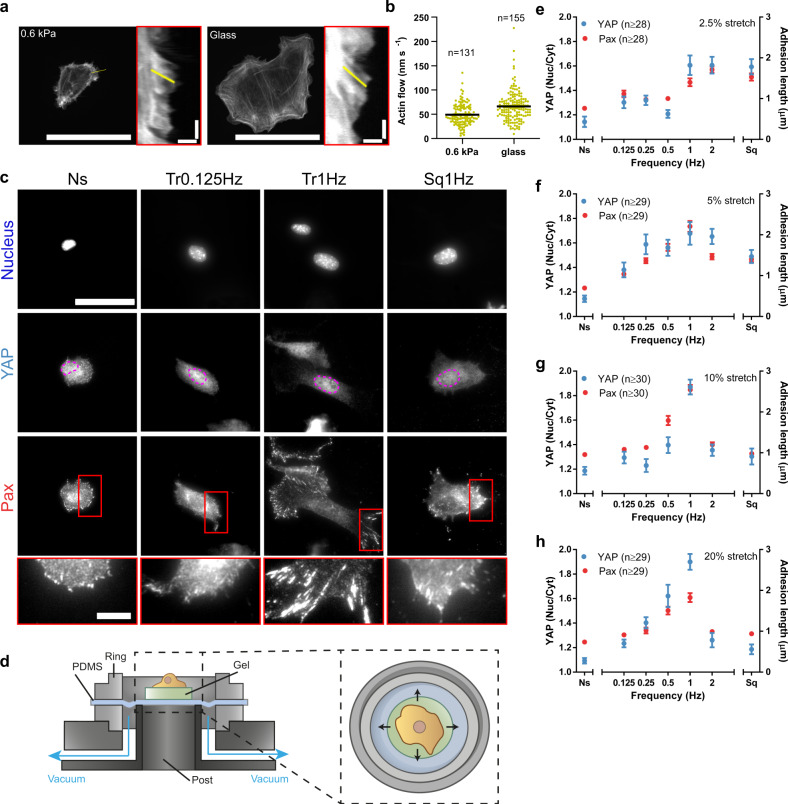
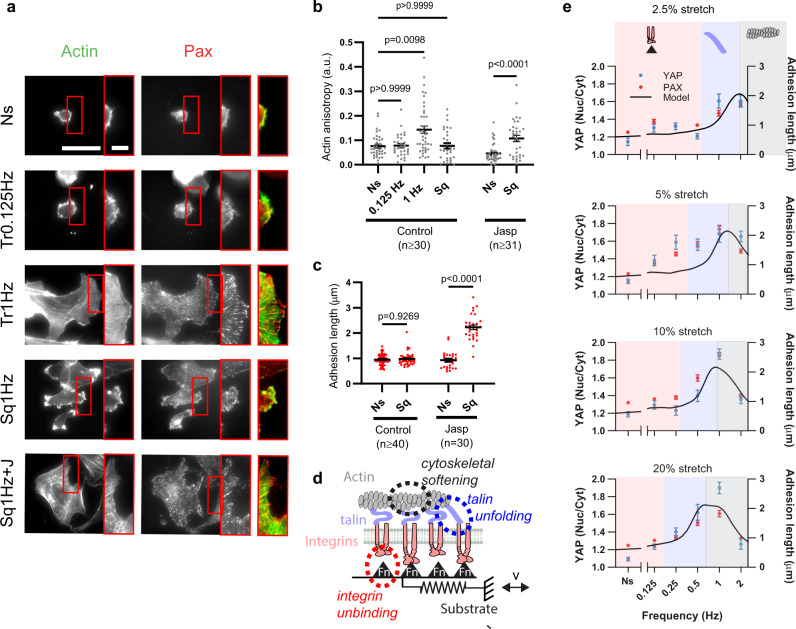
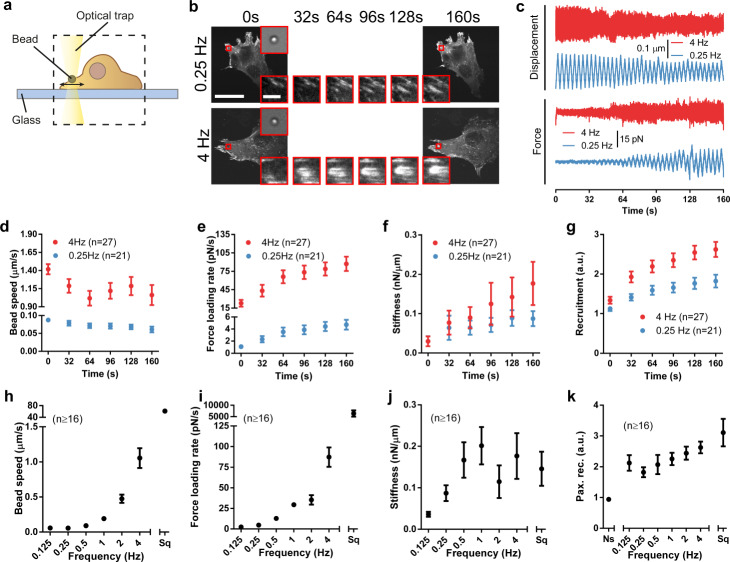
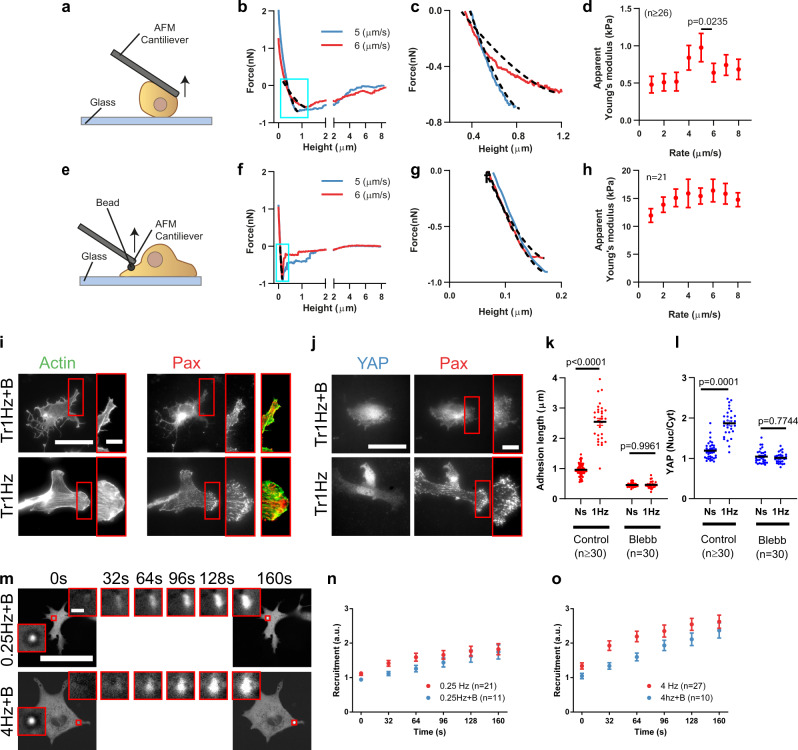
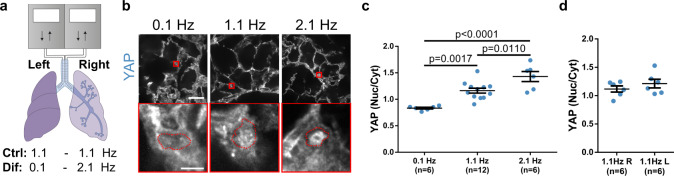
Cell response to force regulates essential processes in health and disease. However, the fundamental mechanical variables that cells sense and respond to remain unclear. Here we show that the rate of force application (loading rate) drives mechanosensing, as predicted by a molecular clutch model. By applying dynamic force regimes to cells through substrate stretching, optical tweezers, and atomic force microscopy, we find that increasing loading rates trigger talin-dependent mechanosensing, leading to adhesion growth and reinforcement, and YAP nuclear localization. However, above a given threshold the actin cytoskeleton softens, decreasing loading rates and preventing reinforcement. By stretching rat lungs in vivo, we show that a similar phenomenon may occur. Our results show that cell sensing of external forces and of passive mechanical parameters (like tissue stiffness) can be understood through the same mechanisms, driven by the properties under force of the mechanosensing molecules involved.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures