

Caenorhabditis elegans RMI2 functional homolog-2 (RMIF-2) and RMI1 (RMH-1) have both overlapping and distinct meiotic functions within the BTR complex
- PMID: 34252074
- PMCID: PMC8318279
- DOI: 10.1371/journal.pgen.1009663
Caenorhabditis elegans RMI2 functional homolog-2 (RMIF-2) and RMI1 (RMH-1) have both overlapping and distinct meiotic functions within the BTR complex
Abstract
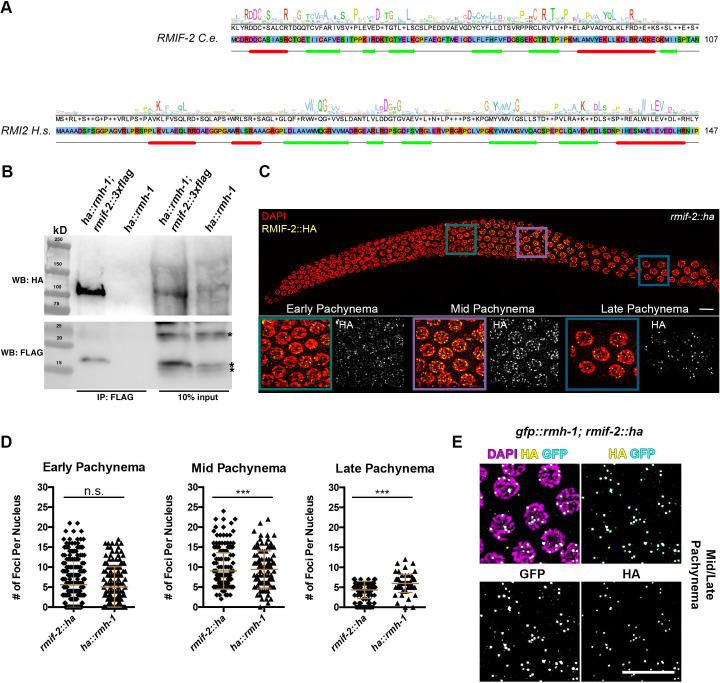
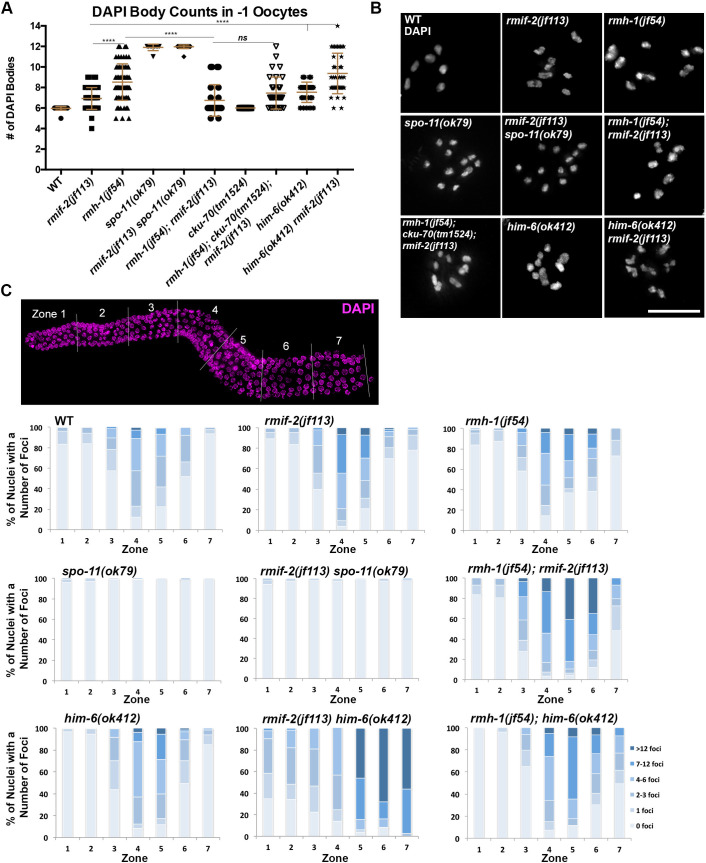
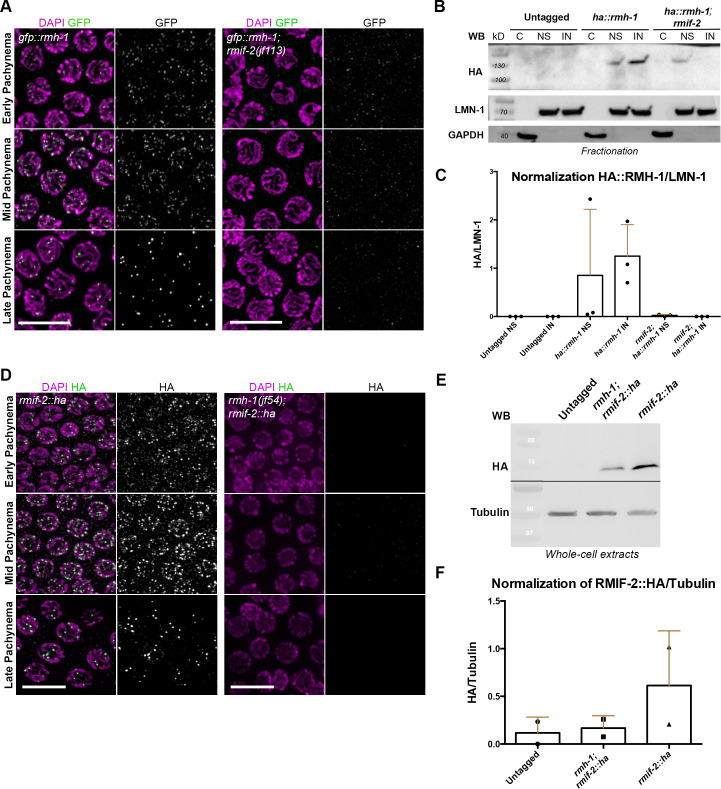
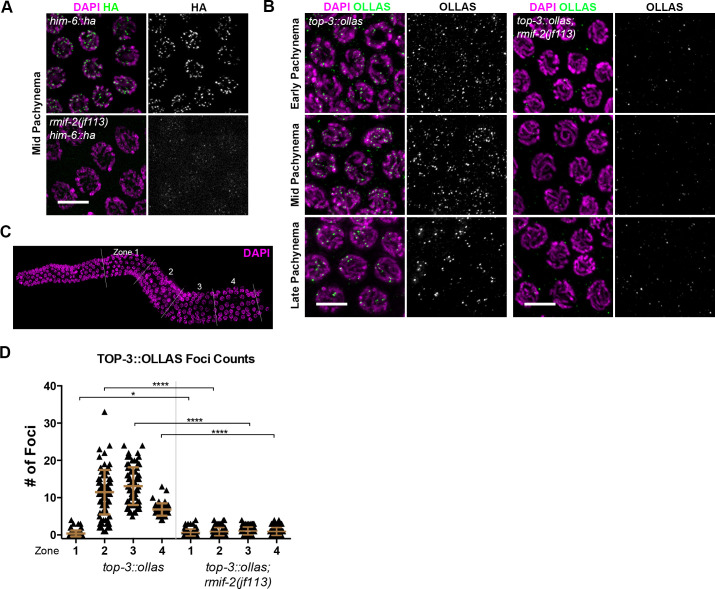
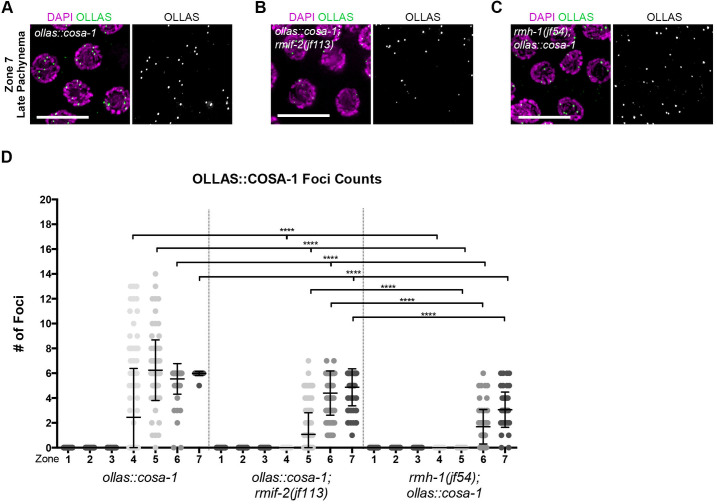
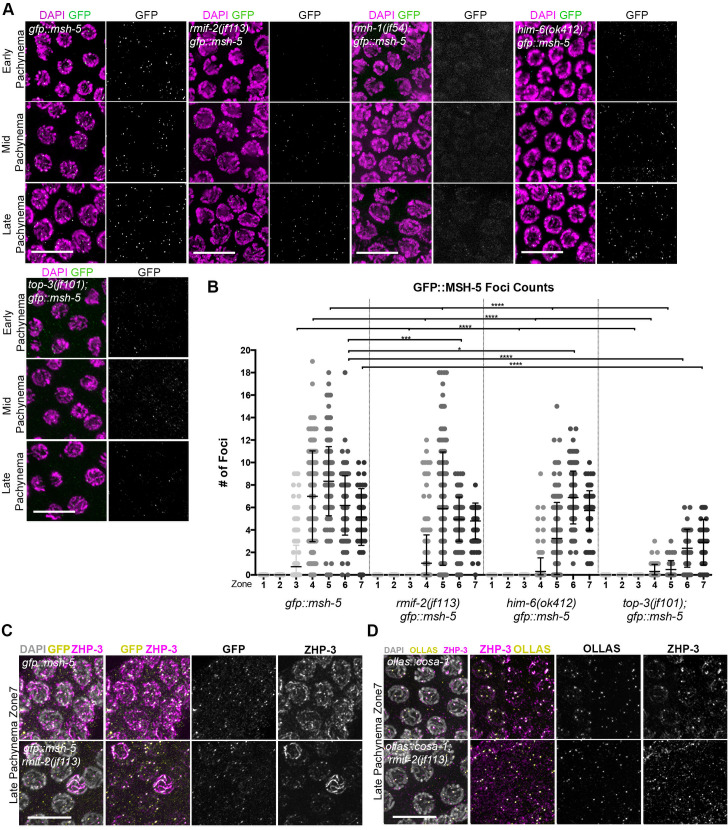
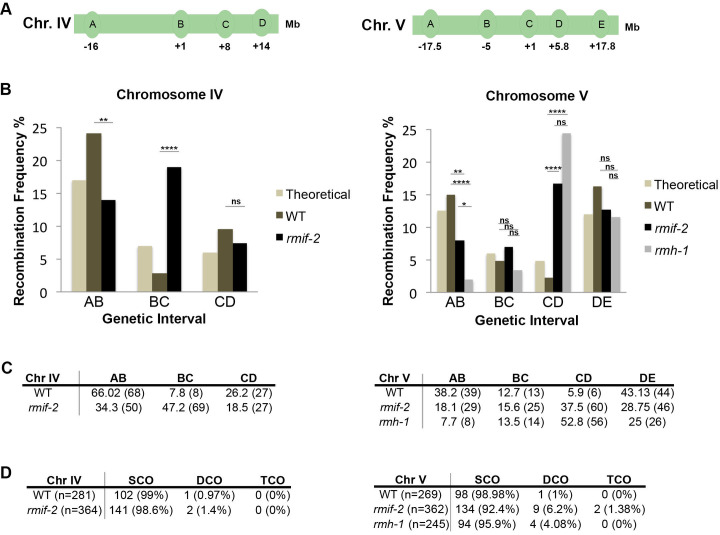
Homologous recombination is a high-fidelity repair pathway for DNA double-strand breaks employed during both mitotic and meiotic cell divisions. Such repair can lead to genetic exchange, originating from crossover (CO) generation. In mitosis, COs are suppressed to prevent sister chromatid exchange. Here, the BTR complex, consisting of the Bloom helicase (HIM-6 in worms), topoisomerase 3 (TOP-3), and the RMI1 (RMH-1 and RMH-2) and RMI2 scaffolding proteins, is essential for dismantling joint DNA molecules to form non-crossovers (NCOs) via decatenation. In contrast, in meiosis COs are essential for accurate chromosome segregation and the BTR complex plays distinct roles in CO and NCO generation at different steps in meiotic recombination. RMI2 stabilizes the RMI1 scaffolding protein, and lack of RMI2 in mitosis leads to elevated sister chromatid exchange, as observed upon RMI1 knockdown. However, much less is known about the involvement of RMI2 in meiotic recombination. So far, RMI2 homologs have been found in vertebrates and plants, but not in lower organisms such as Drosophila, yeast, or worms. We report the identification of the Caenorhabditis elegans functional homolog of RMI2, which we named RMIF-2. The protein shows a dynamic localization pattern to recombination foci during meiotic prophase I and concentration into recombination foci is mutually dependent on other BTR complex proteins. Comparative analysis of the rmif-2 and rmh-1 phenotypes revealed numerous commonalities, including in regulating CO formation and directing COs toward chromosome arms. Surprisingly, the prevalence of heterologous recombination was several fold lower in the rmif-2 mutant, suggesting that RMIF-2 may be dispensable or less strictly required for some BTR complex-mediated activities during meiosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
