

PCSK9 deficiency rewires heart metabolism and drives heart failure with preserved ejection fraction
- PMID: 34252181
- PMCID: PMC8380058
- DOI: 10.1093/eurheartj/ehab431
PCSK9 deficiency rewires heart metabolism and drives heart failure with preserved ejection fraction
Abstract
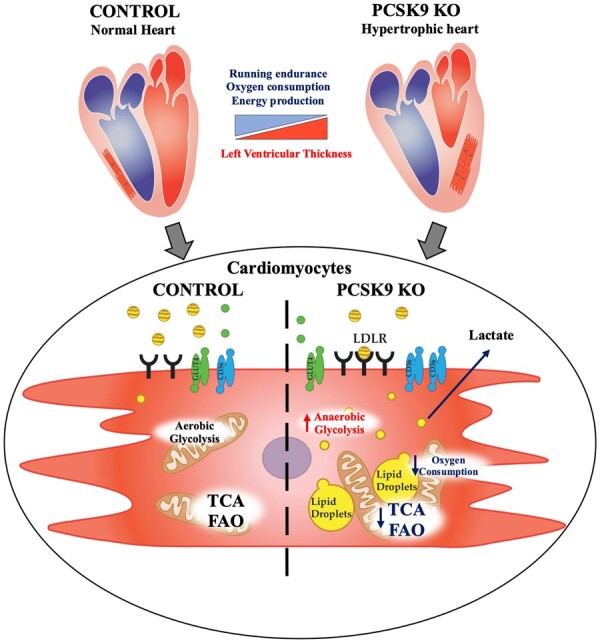
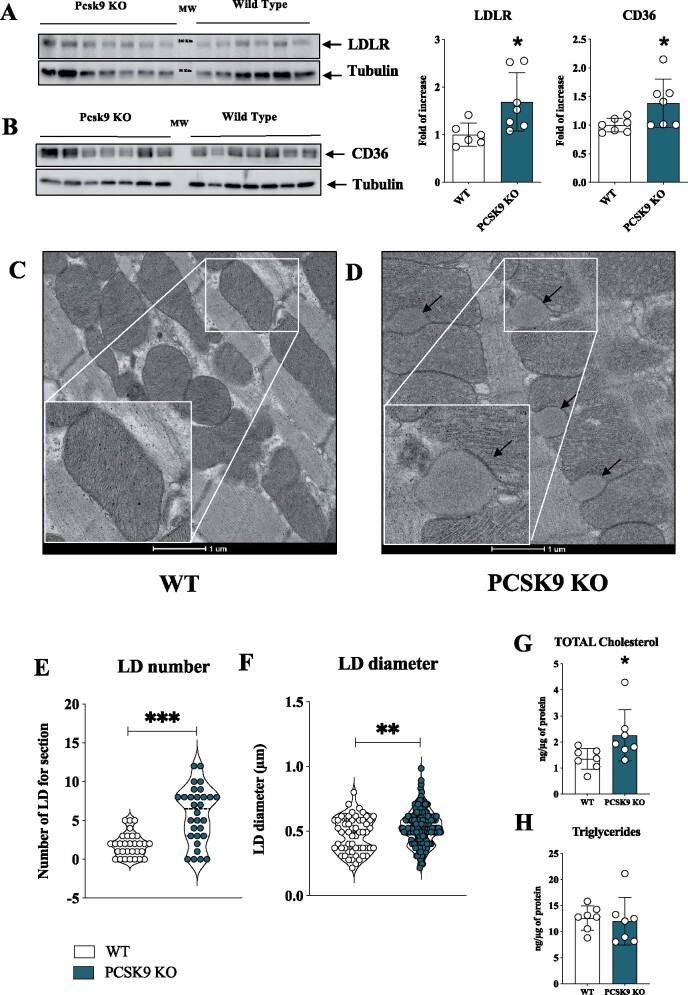
Aims: PCSK9 is secreted into the circulation, mainly by the liver, and interacts with low-density lipoprotein receptor (LDLR) homologous and non-homologous receptors, including CD36, thus favouring their intracellular degradation. As PCSK9 deficiency increases the expression of lipids and lipoprotein receptors, thus contributing to cellular lipid accumulation, we investigated whether this could affect heart metabolism and function.
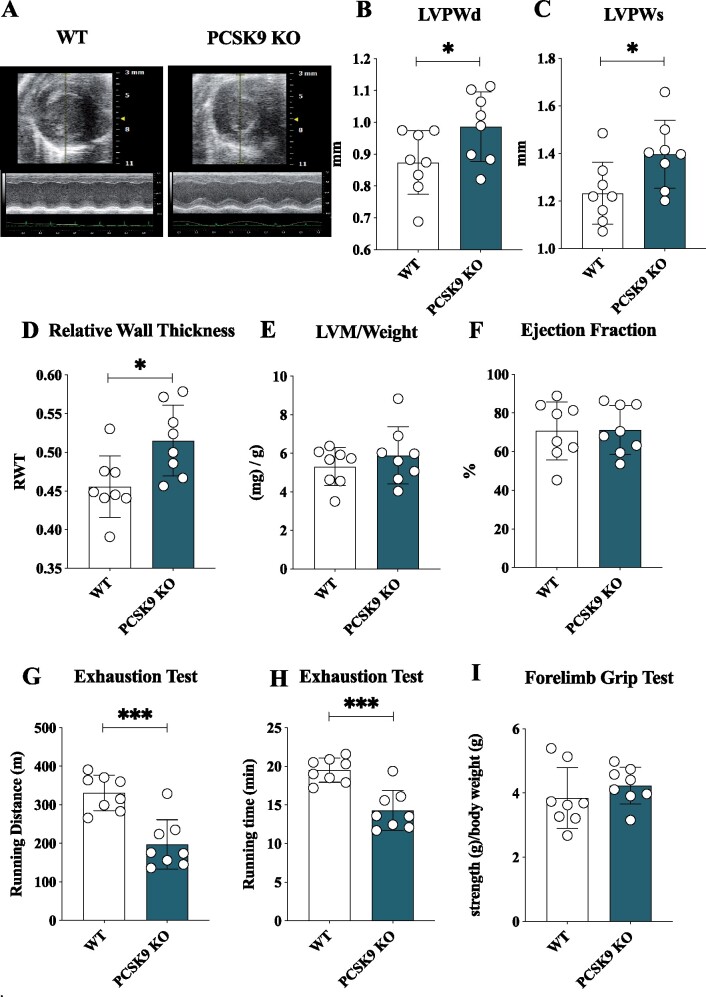
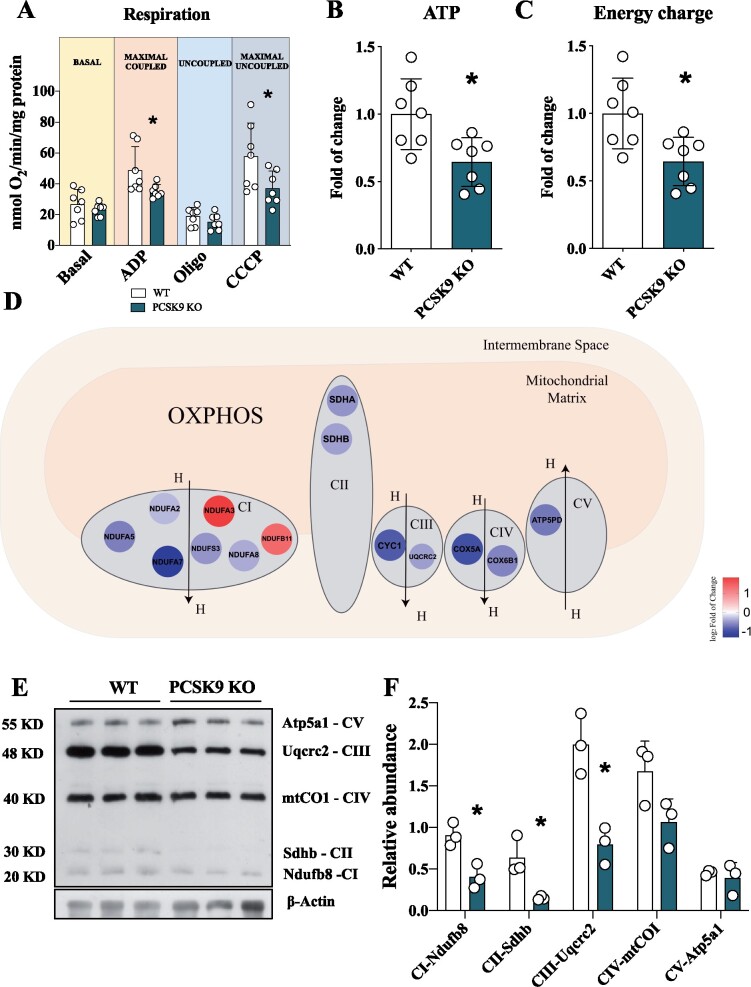
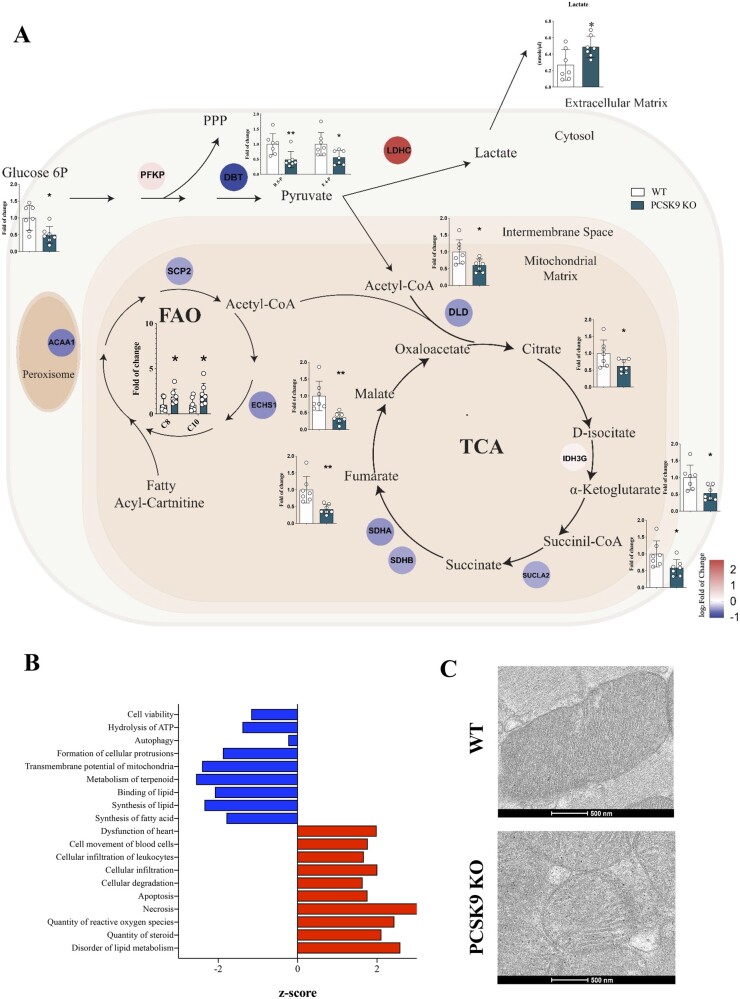
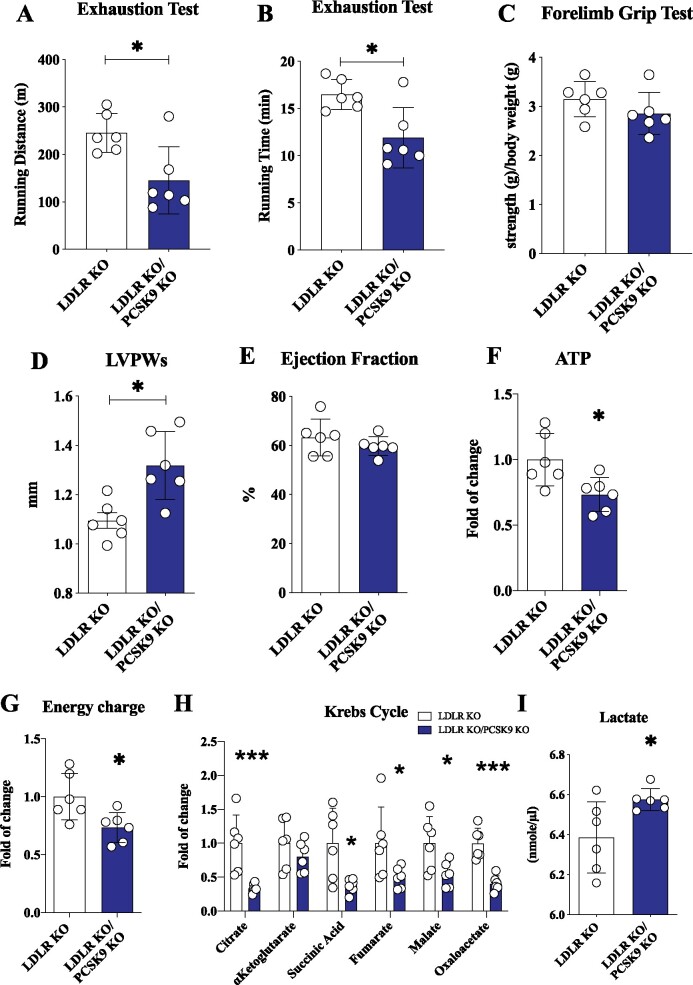
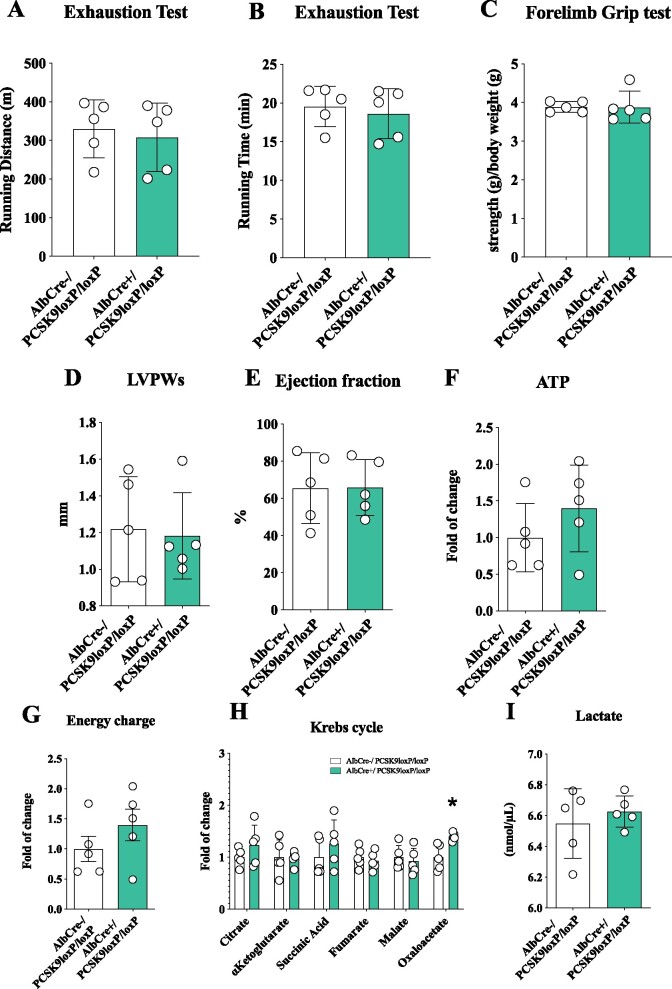
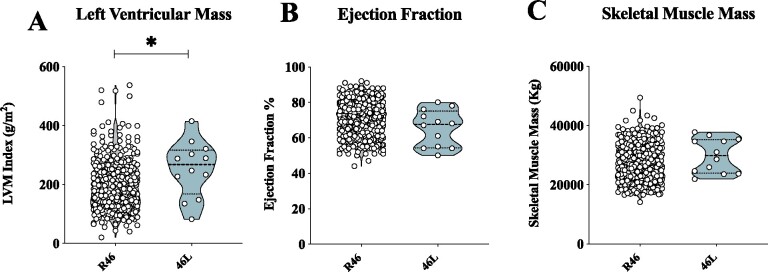
Methods and results: Wild-type (WT), Pcsk9 KO, Liver conditional Pcsk9 KO and Pcsk9/Ldlr double KO male mice were fed for 20 weeks with a standard fat diet and then exercise resistance, muscle strength, and heart characteristics were evaluated. Pcsk9 KO presented reduced running resistance coupled to echocardiographic abnormalities suggestive of heart failure with preserved ejection fraction (HFpEF). Heart mitochondrial activity, following maximal coupled and uncoupled respiration, was reduced in Pcsk9 KO mice compared to WT mice and was coupled to major changes in cardiac metabolism together with increased expression of LDLR and CD36 and with lipid accumulation. A similar phenotype was observed in Pcsk9/Ldlr DKO, thus excluding a contribution for LDLR to cardiac impairment observed in Pcsk9 KO mice. Heart function profiling of the liver selective Pcsk9 KO model further excluded the involvement of circulating PCSK9 in the development of HFpEF, pointing to a possible role locally produced PCSK9. Concordantly, carriers of the R46L loss-of-function variant for PCSK9 presented increased left ventricular mass but similar ejection fraction compared to matched control subjects.
Conclusion: PCSK9 deficiency impacts cardiac lipid metabolism in an LDLR independent manner and contributes to the development of HFpEF.
Keywords: Cholesterol; HFpEF; Heart; LDLR; PCSK9.
© The Author(s) 2021. Published by Oxford University Press on behalf of the European Society of Cardiology.
Figures
Comment in
-
Heart to heart with PCSK9.Eur Heart J. 2021 Aug 21;42(32):3091-3093. doi: 10.1093/eurheartj/ehab480. Eur Heart J. 2021. PMID: 34324660 No abstract available.
References
-
- Stanley WC, Recchia FA, Lopaschuk GD. Myocardial substrate metabolism in the normal and failing heart. Physiol Rev 2005;85:1093–1129. - PubMed
-
- Pulinilkunnil T, Rodrigues B. Cardiac lipoprotein lipase: metabolic basis for diabetic heart disease. Cardiovasc Res 2006;69:329–340. - PubMed
-
- van der Vusse GJ, van Bilsen M, Glatz JF. Cardiac fatty acid uptake and transport in health and disease. Cardiovasc Res 2000;45:279–293. - PubMed
-
- Bharadwaj KG, Hiyama Y, Hu Y, Huggins LA, Ramakrishnan R, Abumrad NA, Shulman GI, Blaner WS, Goldberg IJ. Chylomicron- and VLDL-derived lipids enter the heart through different pathways: in vivo evidence for receptor- and non-receptor-mediated fatty acid uptake. J Biol Chem 2010;285:37976–37986. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
