

Diversity and function of arthropod endosymbiont toxins
- PMID: 34253453
- PMCID: PMC8742837
- DOI: 10.1016/j.tim.2021.06.008
Diversity and function of arthropod endosymbiont toxins
Abstract
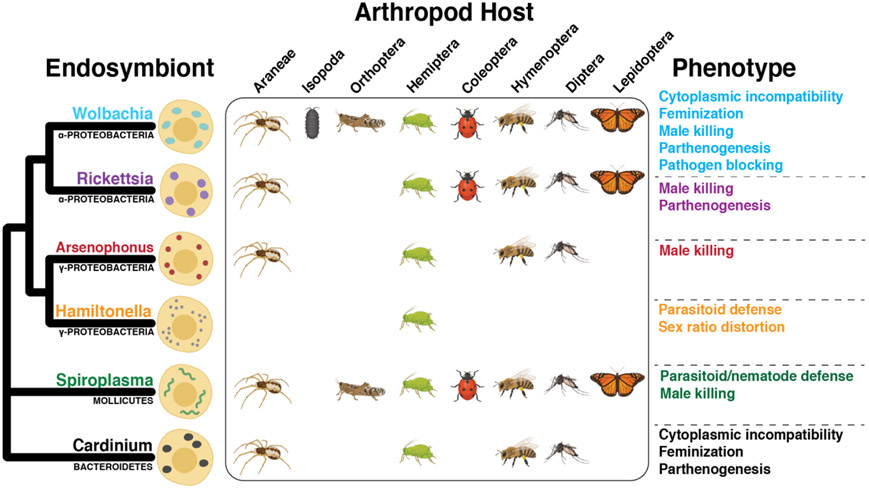
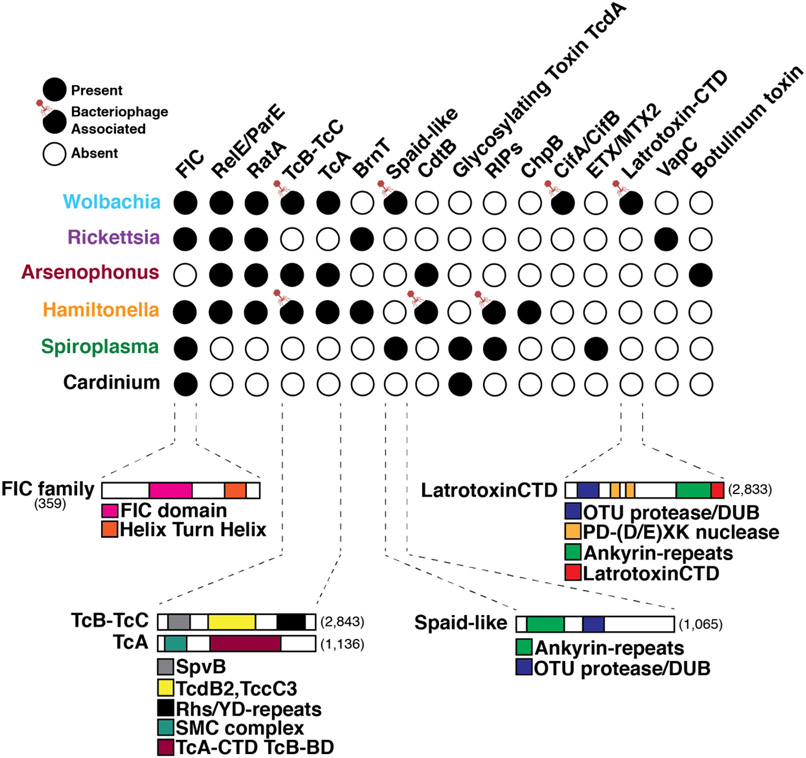
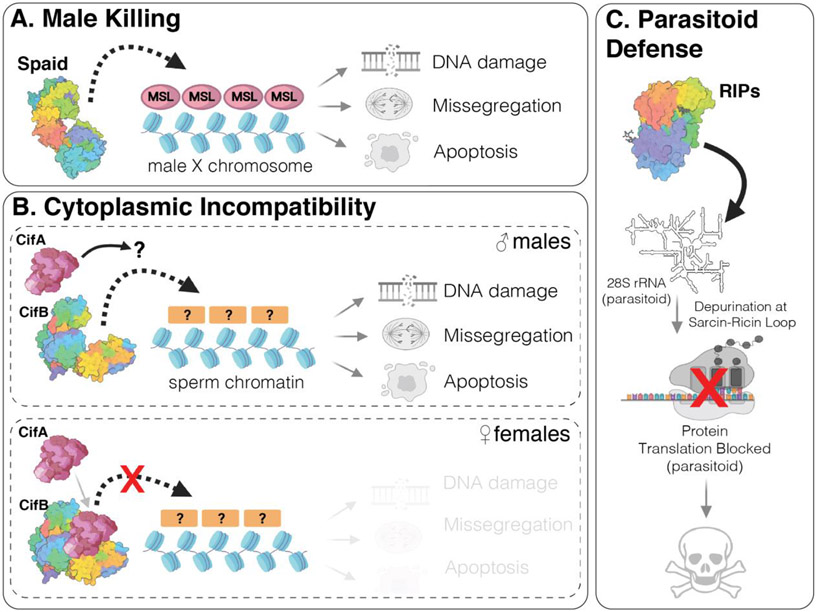
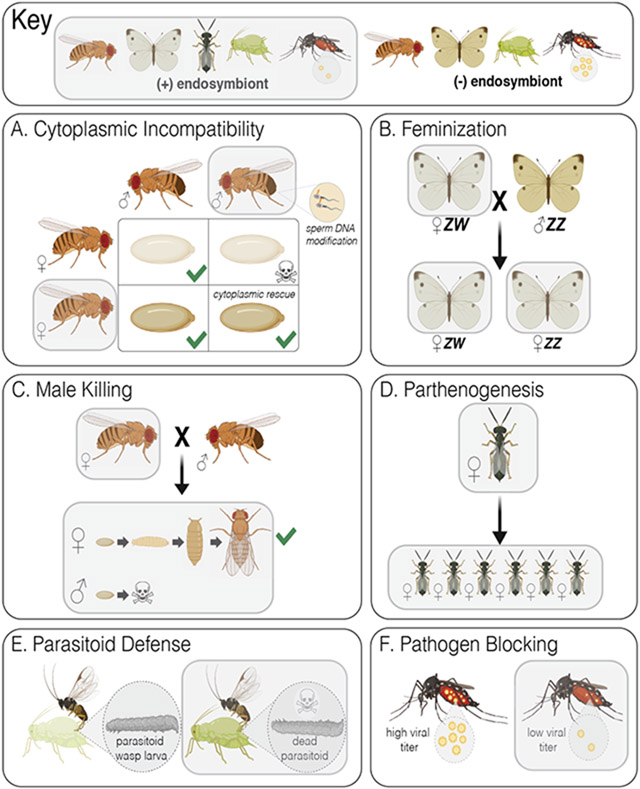
Bacterial endosymbionts induce dramatic phenotypes in their arthropod hosts, including cytoplasmic incompatibility, feminization, parthenogenesis, male killing, parasitoid defense, and pathogen blocking. The molecular mechanisms underlying these effects remain largely unknown but recent evidence suggests that protein toxins secreted by the endosymbionts play a role. Here, we describe the diversity and function of endosymbiont proteins with homology to known bacterial toxins. We focus on maternally transmitted endosymbionts belonging to the Wolbachia, Rickettsia, Arsenophonus, Hamiltonella, Spiroplasma, and Cardinium genera because of their ability to induce the above phenotypes. We identify at least 16 distinct toxin families with diverse enzymatic activities, including AMPylases, nucleases, proteases, and glycosyltransferases. Notably, several annotated toxins contain domains with homology to eukaryotic proteins, suggesting that arthropod endosymbionts mimic host biochemistry to manipulate host physiology, similar to bacterial pathogens.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests There are no interests to declare.
Figures
Similar articles
-
Low Endosymbiont Incidence in Drosophila Species Across Peninsula Thailand.Microb Ecol. 2023 Feb;85(2):730-736. doi: 10.1007/s00248-022-01982-1. Epub 2022 Feb 22. Microb Ecol. 2023. PMID: 35192040
-
Infections with Wolbachia, Spiroplasma, and Rickettsia in the Dolichopodidae and other Empidoidea.Infect Genet Evol. 2013 Jan;13:317-30. doi: 10.1016/j.meegid.2012.11.005. Epub 2012 Nov 24. Infect Genet Evol. 2013. PMID: 23183308
-
Rickettsia induces strong cytoplasmic incompatibility in a predatory insect.Proc Biol Sci. 2024 Jul;291(2027):20240680. doi: 10.1098/rspb.2024.0680. Epub 2024 Jul 31. Proc Biol Sci. 2024. PMID: 39079670 Free PMC article.
-
Manipulation of arthropod sex determination by endosymbionts: diversity and molecular mechanisms.Sex Dev. 2014;8(1-3):59-73. doi: 10.1159/000357024. Epub 2013 Dec 20. Sex Dev. 2014. PMID: 24355929 Review.
-
Molecular Biology of Cytoplasmic Incompatibility Caused by Wolbachia Endosymbionts.Annu Rev Microbiol. 2023 Sep 15;77:299-316. doi: 10.1146/annurev-micro-041020-024616. Epub 2023 Jun 7. Annu Rev Microbiol. 2023. PMID: 37285552 Review.
Cited by
-
Microbial diversity of ticks and a novel typhus group Rickettsia species (Rickettsiales bacterium Ac37b) in Inner Mongolia, China.Parasite. 2023;30:58. doi: 10.1051/parasite/2023057. Epub 2023 Dec 12. Parasite. 2023. PMID: 38084939 Free PMC article.
-
Metagenome diversity illuminates the origins of pathogen effectors.mBio. 2024 May 8;15(5):e0075923. doi: 10.1128/mbio.00759-23. Epub 2024 Apr 2. mBio. 2024. PMID: 38564675 Free PMC article.
-
Phylogenomic analysis of Wolbachia genomes from the Darwin Tree of Life biodiversity genomics project.PLoS Biol. 2023 Jan 23;21(1):e3001972. doi: 10.1371/journal.pbio.3001972. eCollection 2023 Jan. PLoS Biol. 2023. PMID: 36689552 Free PMC article.
-
'Candidatus Megaira' are diverse symbionts of algae and ciliates with the potential for defensive symbiosis.Microb Genom. 2023 Mar;9(3):mgen000950. doi: 10.1099/mgen.0.000950. Microb Genom. 2023. PMID: 36897930 Free PMC article.
-
Insect-microbe interactions and their influence on organisms and ecosystems.Ecol Evol. 2024 Jul 21;14(7):e11699. doi: 10.1002/ece3.11699. eCollection 2024 Jul. Ecol Evol. 2024. PMID: 39041011 Free PMC article. Review.
References
-
- Giribet G and Edgecombe GD (2012) Reevaluating the arthropod tree of life. Annu. Rev. Entomol 57, 167–186 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
