

CD8+ T cell differentiation and dysfunction in cancer
- PMID: 34253904
- PMCID: PMC9792152
- DOI: 10.1038/s41577-021-00574-3
CD8+ T cell differentiation and dysfunction in cancer
Abstract
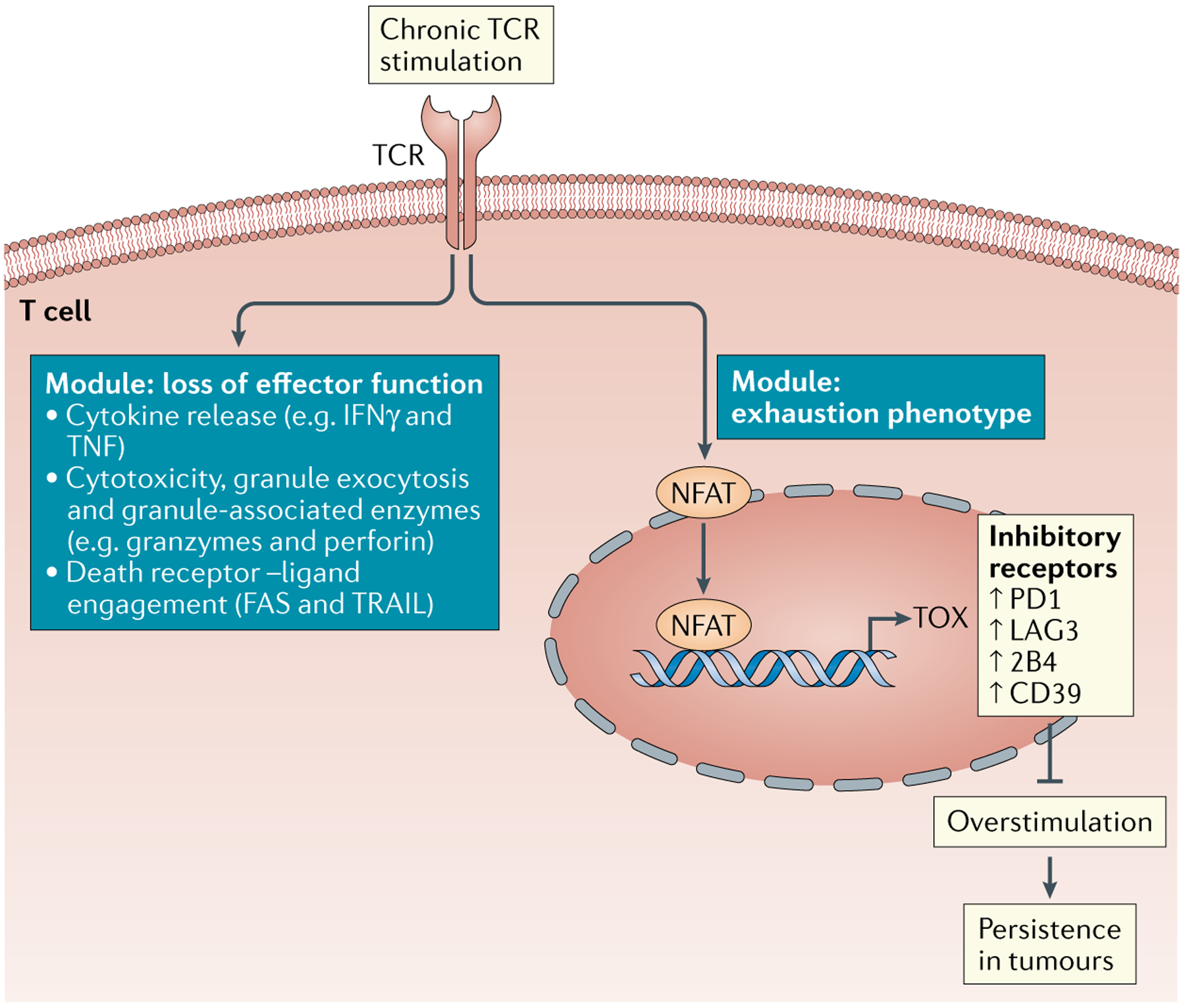
CD8+ T cells specific for cancer cells are detected within tumours. However, despite their presence, tumours progress. The clinical success of immune checkpoint blockade and adoptive T cell therapy demonstrates the potential of CD8+ T cells to mediate antitumour responses; however, most patients with cancer fail to achieve long-term responses to immunotherapy. Here we review CD8+ T cell differentiation to dysfunctional states during tumorigenesis. We highlight similarities and differences between T cell dysfunction and other hyporesponsive T cell states and discuss the spatio-temporal factors contributing to T cell state heterogeneity in tumours. An important challenge is predicting which patients will respond to immunotherapeutic interventions and understanding which T cell subsets mediate the clinical response. We explore our current understanding of what determines T cell responsiveness and resistance to immunotherapy and point out the outstanding research questions.
© 2021. Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures



References
-
- De Plaen E et al. Immunogenic (tum-) variants of mouse tumor P815: cloning of the gene of tum-antigen P91A and identification of the tum- mutation. Proc. Natl Acad. Sci. USA 85, 2274–2278 (1988). - PMC - PubMed
-
This is the first study to identify a somatic mutation in a mouse cancer cell line variant creating a cytolytic T lymphocyte-recognized epitope using a cloning-based approach.
-
- Mandelboim O et al. CTL induction by a tumour-associated antigen octapeptide derived from a murine lung carcinoma. Nature 369, 67–71 (1994). - PubMed
-
This study uses protein extraction and fractionation to identify a peptide epitope in a mouse lung cancer cell line containing an amino acid-altering mutation recognized by cytolytic T lymphocytes.
-
- Monach PA, Meredith SC, Siegel CT & Schreiber H A unique tumor antigen produced by a single amino acid substitution. Immunity 2, 45–59 (1995). - PubMed
-
This study is the first to identify a mutation in a mouse cancer cell creating a tumour-specific CD4+ T cell-recognized epitope.
-
- Wolfel T et al. A p16INK4a-insensitive CDK4 mutant targeted by cytolytic T lymphocytes in a human melanoma. Science 269, 1281–1284 (1995). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
