

The Evolutionary Origins of Extreme Halophilic Archaeal Lineages
- PMID: 34255041
- PMCID: PMC8350355
- DOI: 10.1093/gbe/evab166
The Evolutionary Origins of Extreme Halophilic Archaeal Lineages
Abstract
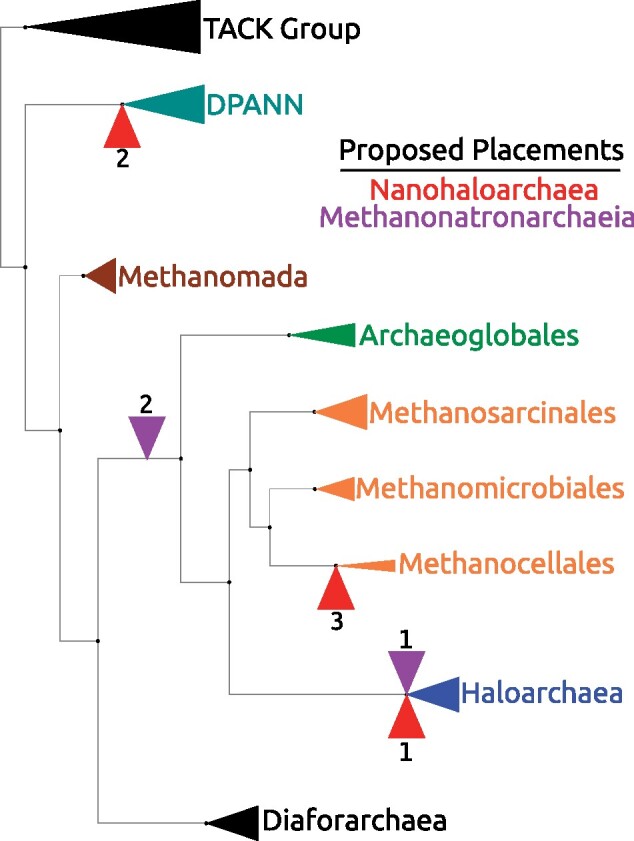
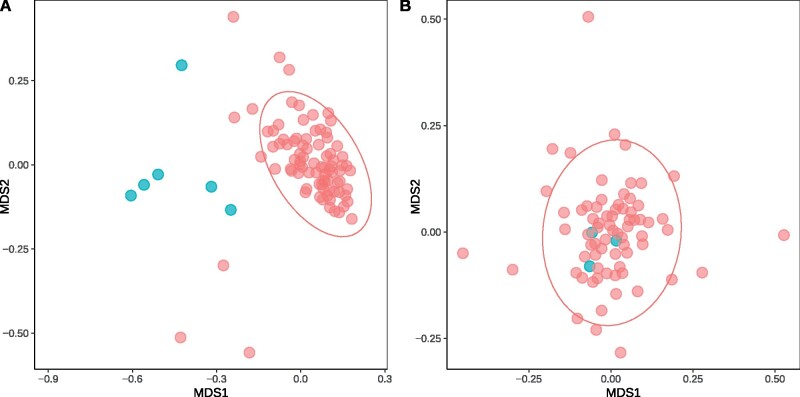
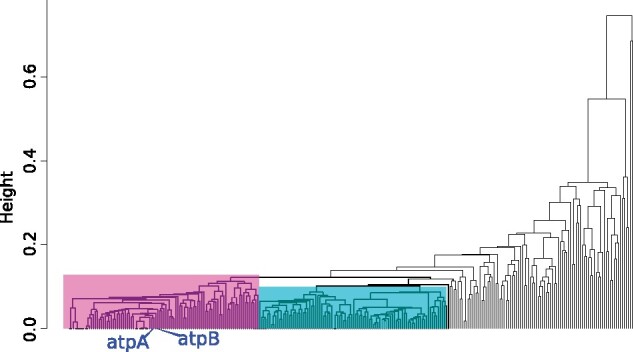
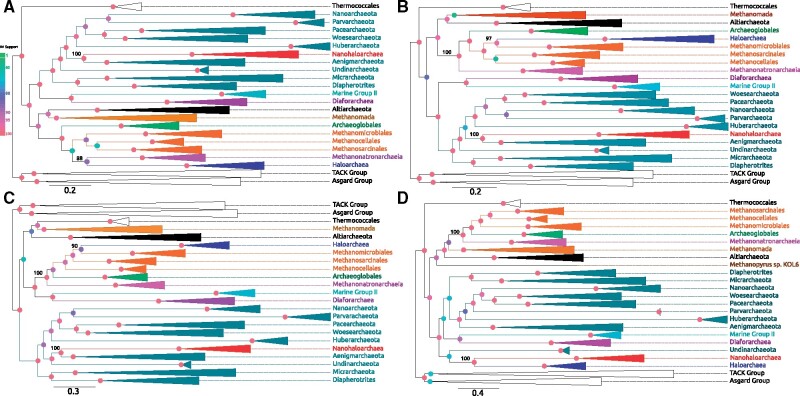
Interest and controversy surrounding the evolutionary origins of extremely halophilic Archaea has increased in recent years, due to the discovery and characterization of the Nanohaloarchaea and the Methanonatronarchaeia. Initial attempts in explaining the evolutionary placement of the two new lineages in relation to the classical Halobacteria (also referred to as Haloarchaea) resulted in hypotheses that imply the new groups share a common ancestor with the Haloarchaea. However, more recent analyses have led to a shift: the Nanohaloarchaea have been largely accepted as being a member of the DPANN superphylum, outside of the euryarchaeota; whereas the Methanonatronarchaeia have been placed near the base of the Methanotecta (composed of the class II methanogens, the Halobacteriales, and Archaeoglobales). These opposing hypotheses have far-reaching implications on the concepts of convergent evolution (distantly related groups evolve similar strategies for survival), genome reduction, and gene transfer. In this work, we attempt to resolve these conflicts with phylogenetic and phylogenomic data. We provide a robust taxonomic sampling of Archaeal genomes that spans the Asgardarchaea, TACK Group, euryarchaeota, and the DPANN superphylum. In addition, we assembled draft genomes from seven new representatives of the Nanohaloarchaea from distinct geographic locations. Phylogenies derived from these data imply that the highly conserved ATP synthase catalytic/noncatalytic subunits of Nanohaloarchaea share a sisterhood relationship with the Haloarchaea. We also employ a novel gene family distance clustering strategy which shows this sisterhood relationship is not likely the result of a recent gene transfer. In addition, we present and evaluate data that argue for and against the monophyly of the DPANN superphylum, in particular, the inclusion of the Nanohaloarchaea in DPANN.
Keywords: Methanonatronarchaeia; Nanohaloarchaea; gene concordance; metagenomic-assembled genome (MAG); single amplified genome (SAG).
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures

References
-
- Ané C, Larget B, Baum DA, Smith SD, Rokas A.. 2007. Bayesian estimation of concordance among gene trees. Mol Biol Evol. 24(7):1575. - PubMed
-
- Aouad M, Borrel G, Brochier-Armanet C, Gribaldo S.. 2019. Evolutionary placement of Methanonatronarchaeia. Nat Microbiol. 4(4):558–559. - PubMed
-
- Aouad M, et al.2018. Extreme halophilic archaea derive from two distinct methanogen Class II lineages. Mol Phylogenet Evol. 127:46–54. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
