

SARS-CoV-2 RBD antibodies that maximize breadth and resistance to escape
- PMID: 34261126
- PMCID: PMC9282883
- DOI: 10.1038/s41586-021-03807-6
SARS-CoV-2 RBD antibodies that maximize breadth and resistance to escape
Abstract
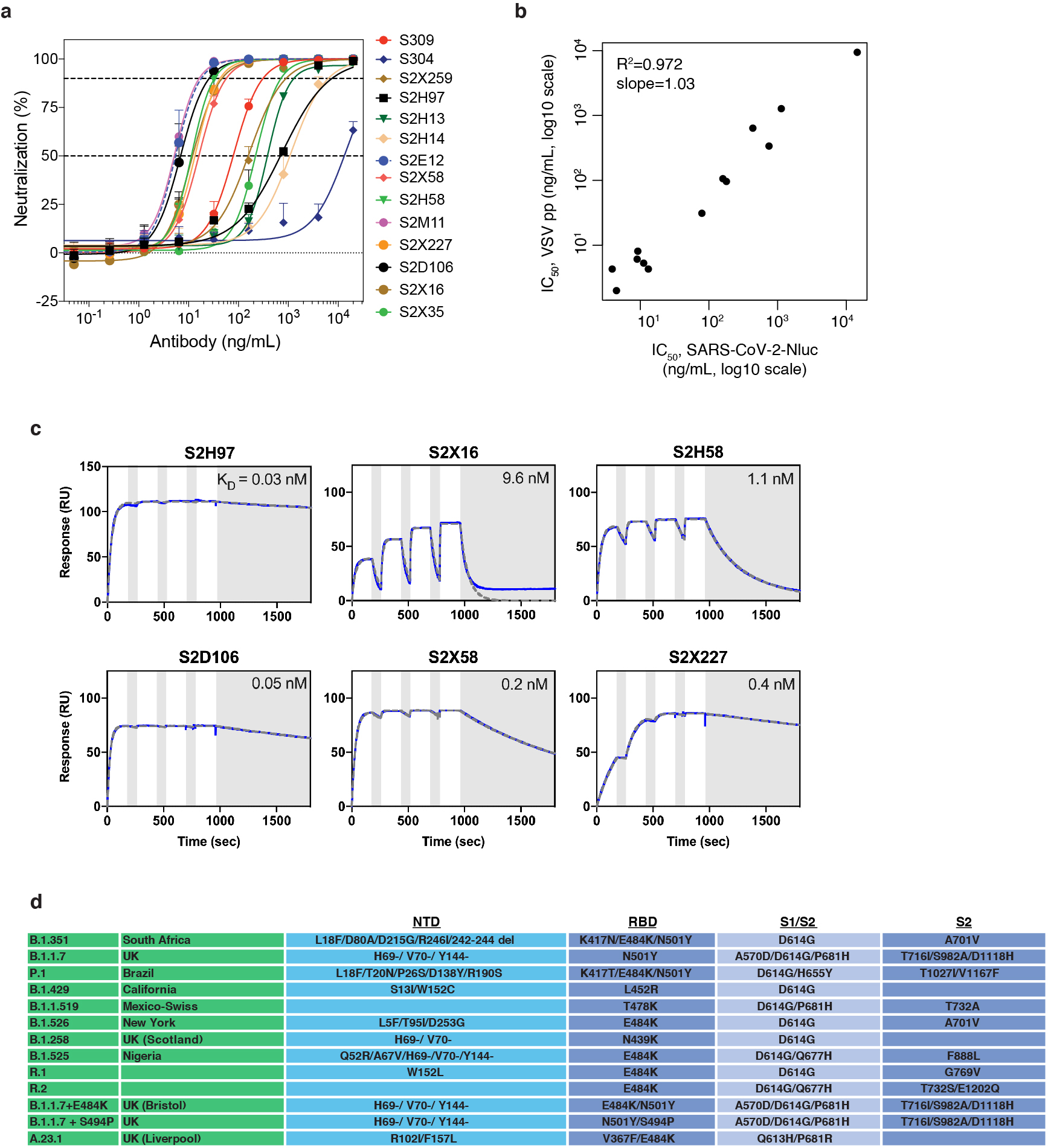
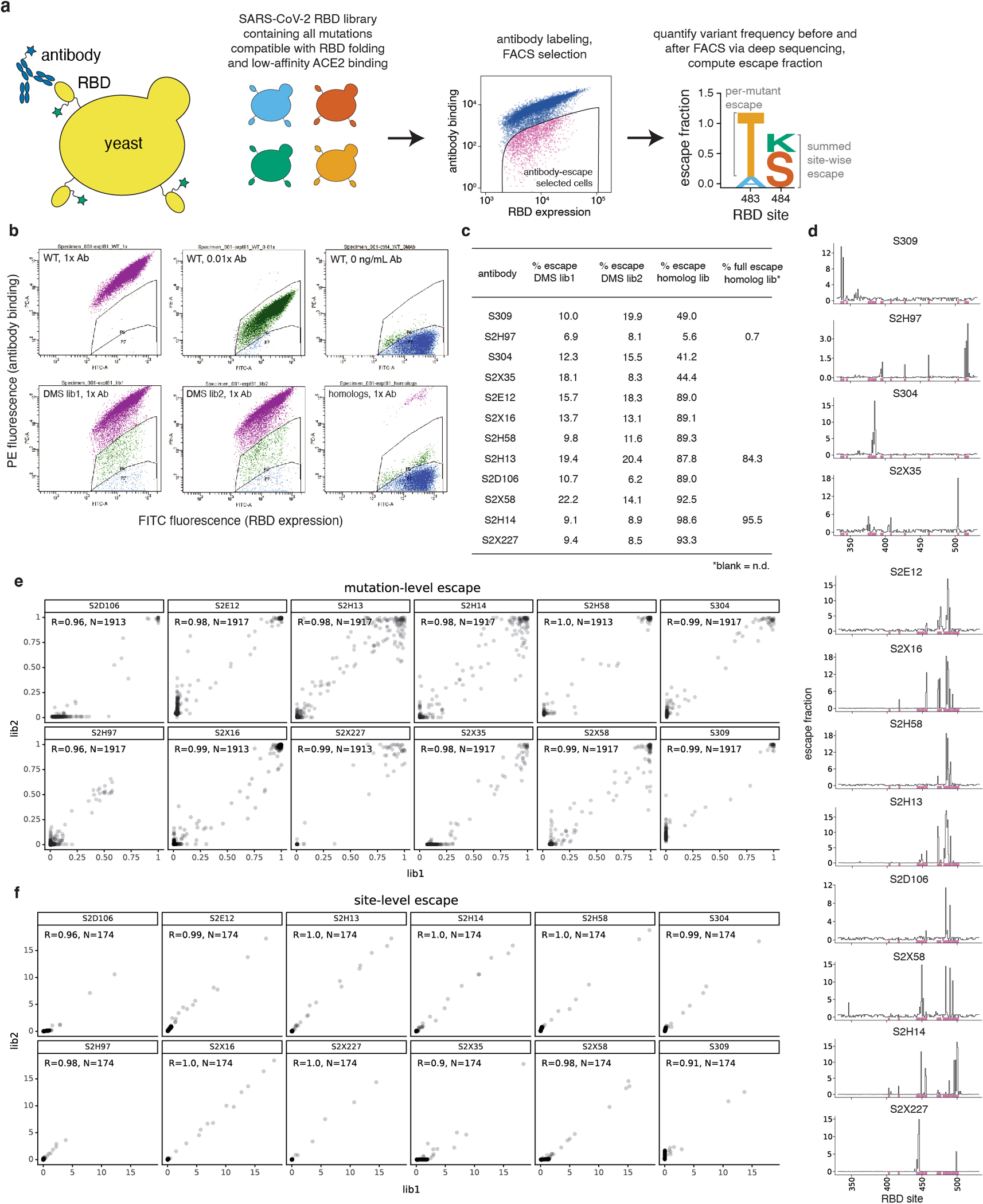
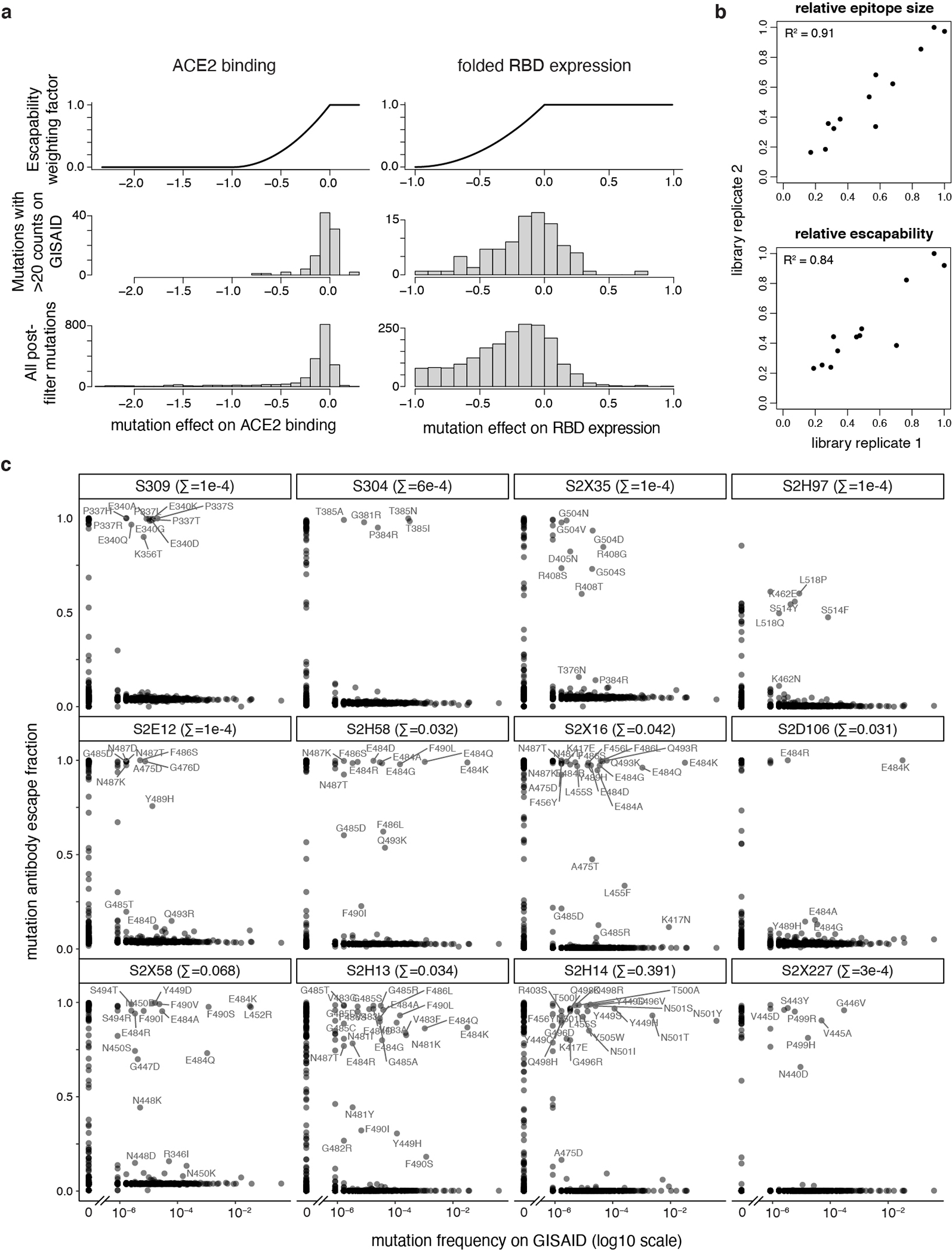
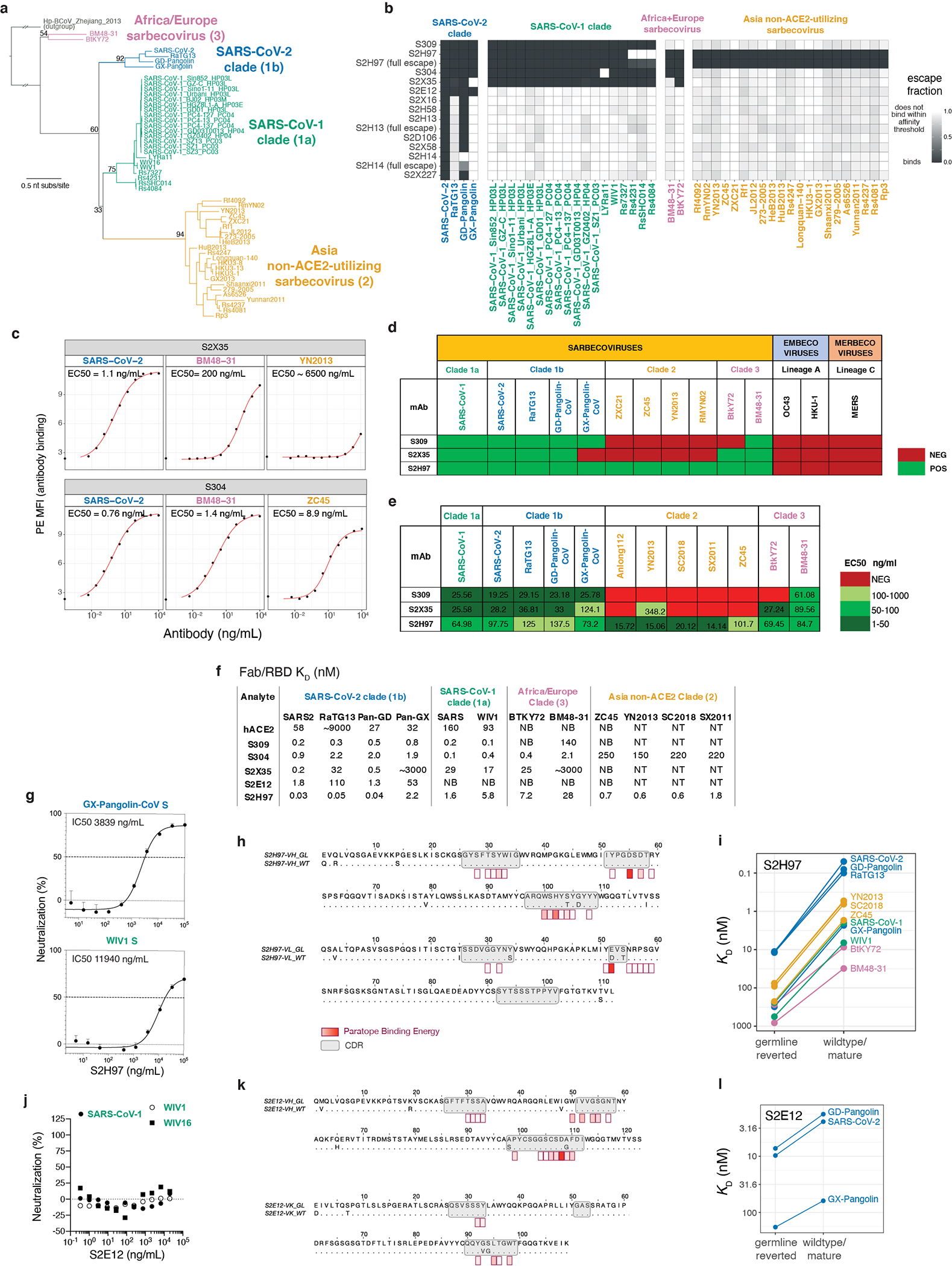
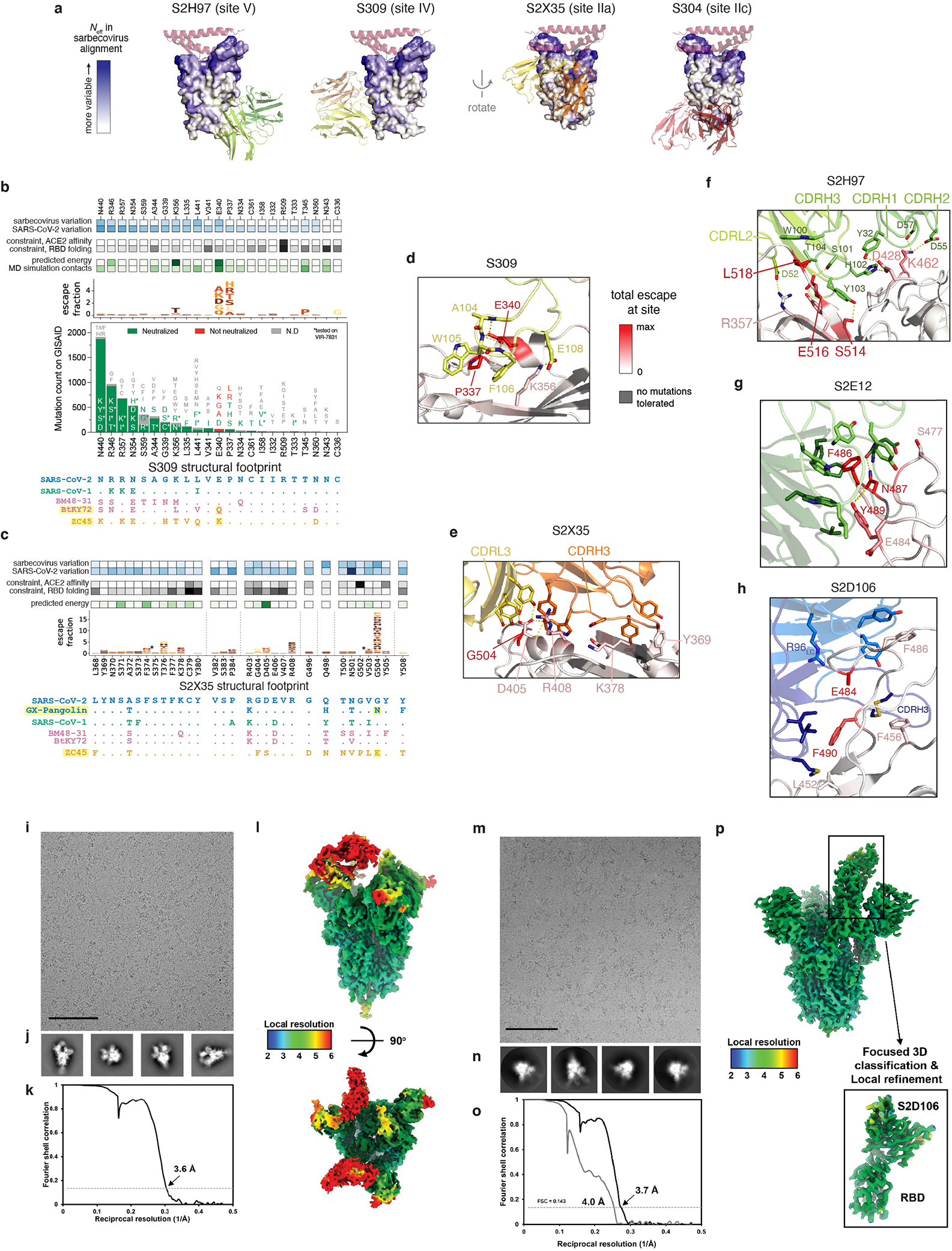
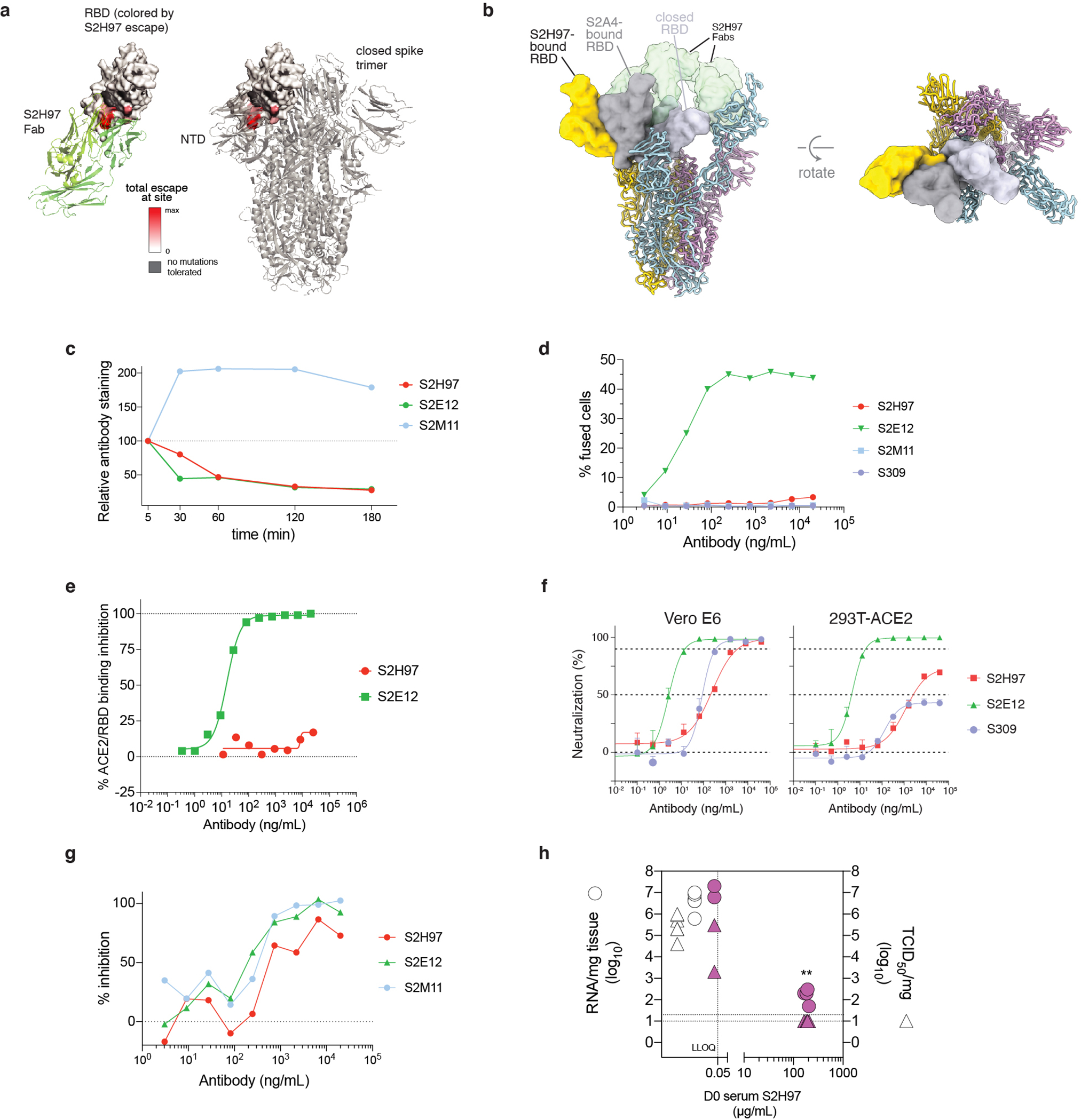
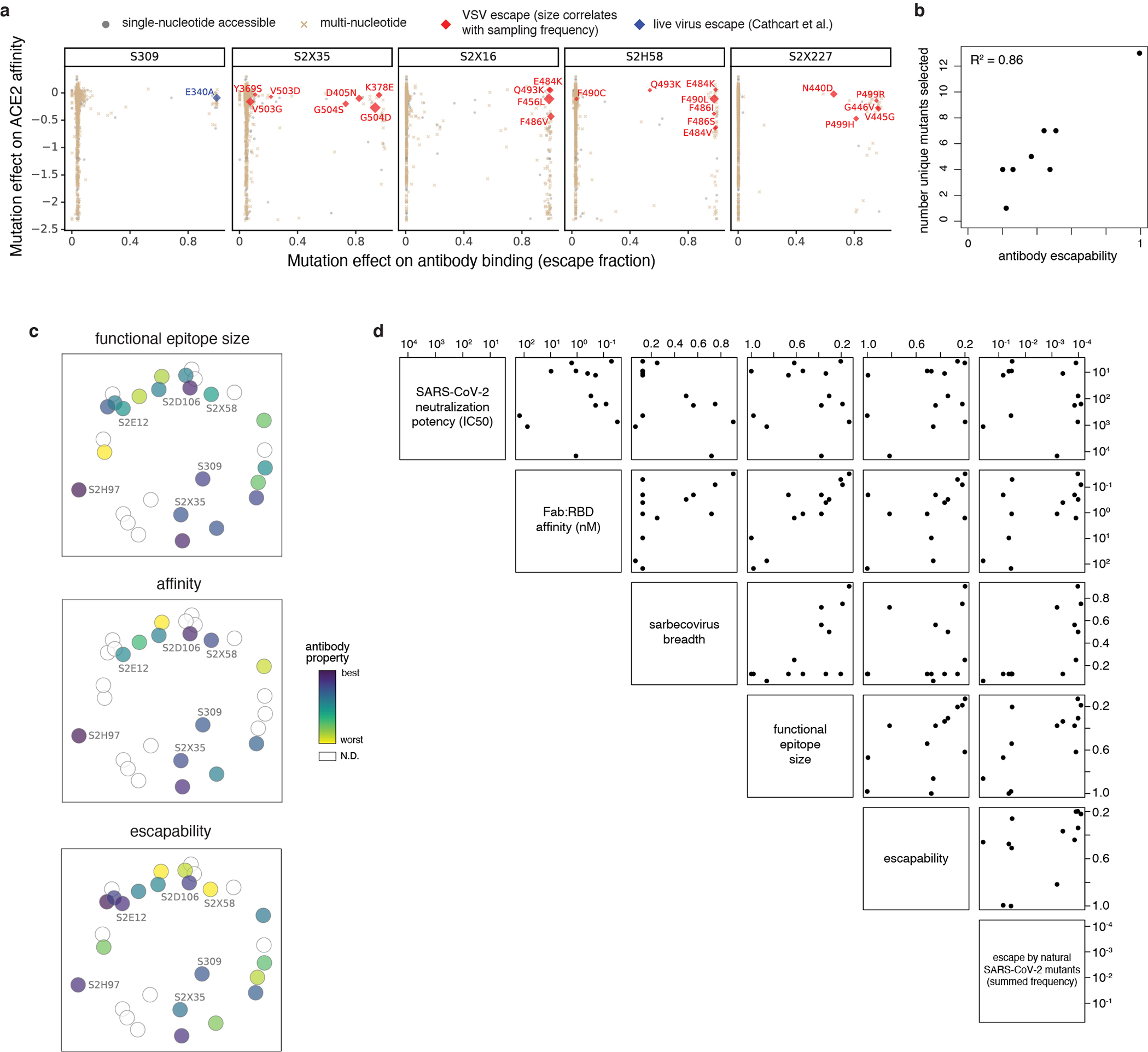
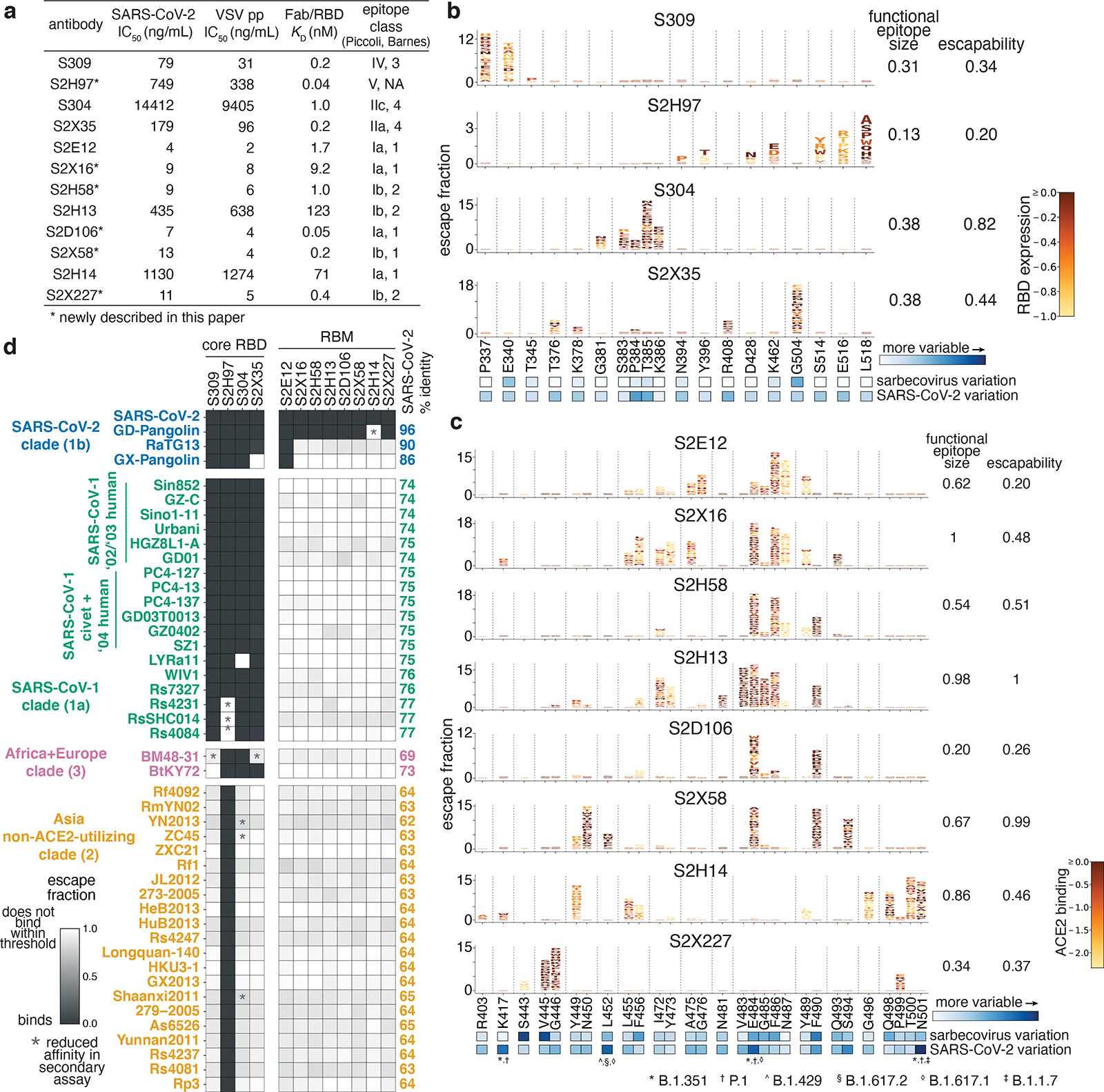
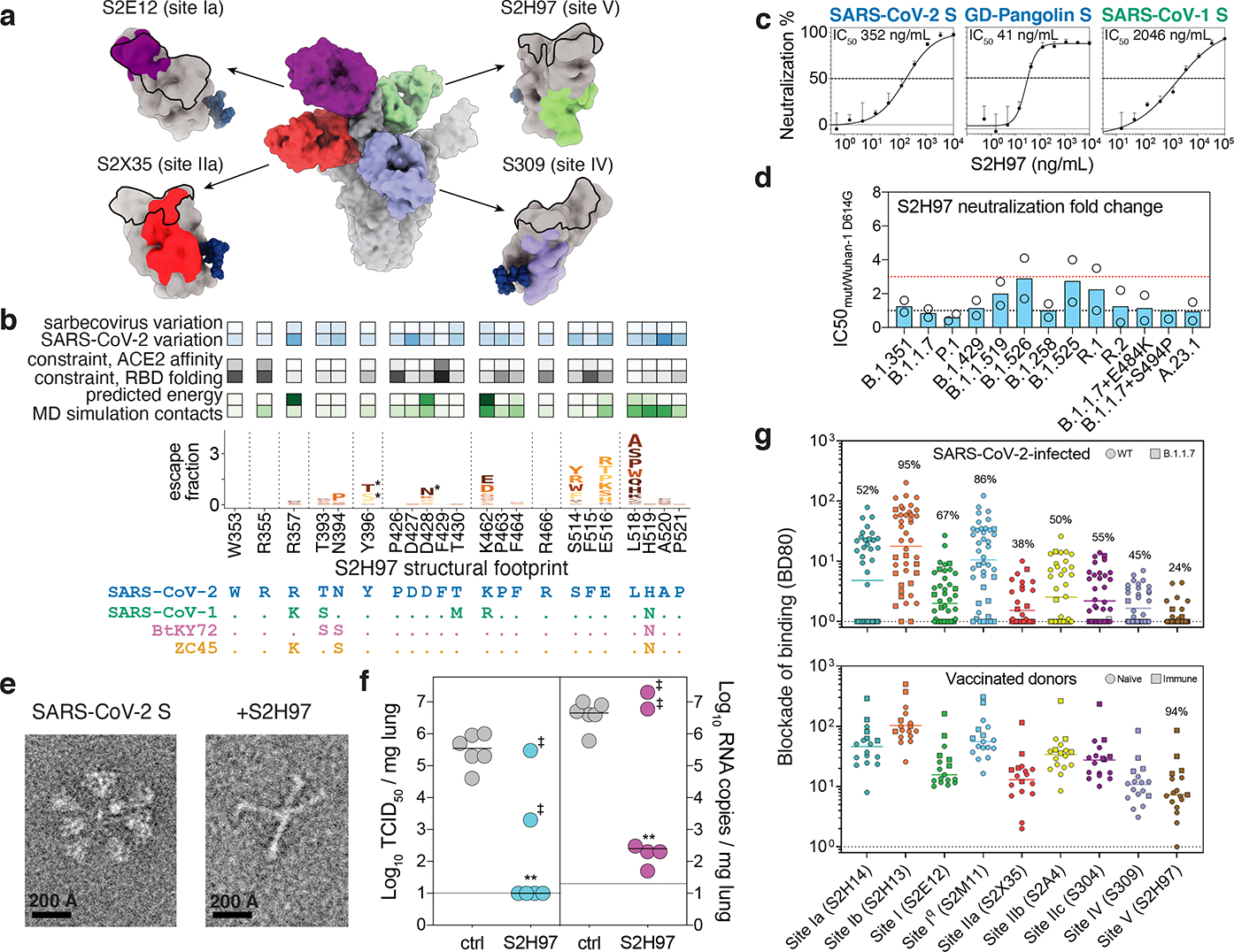
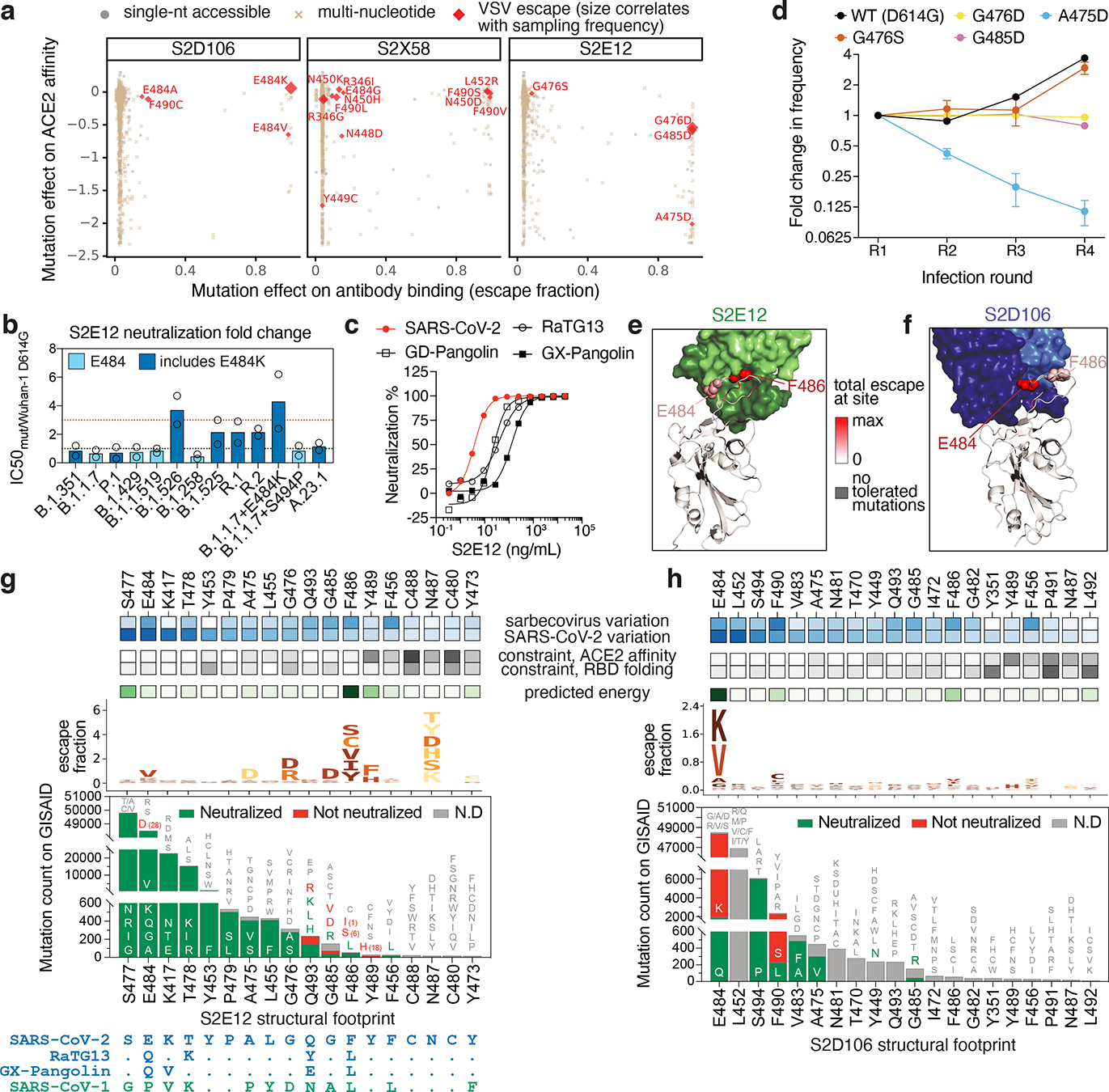
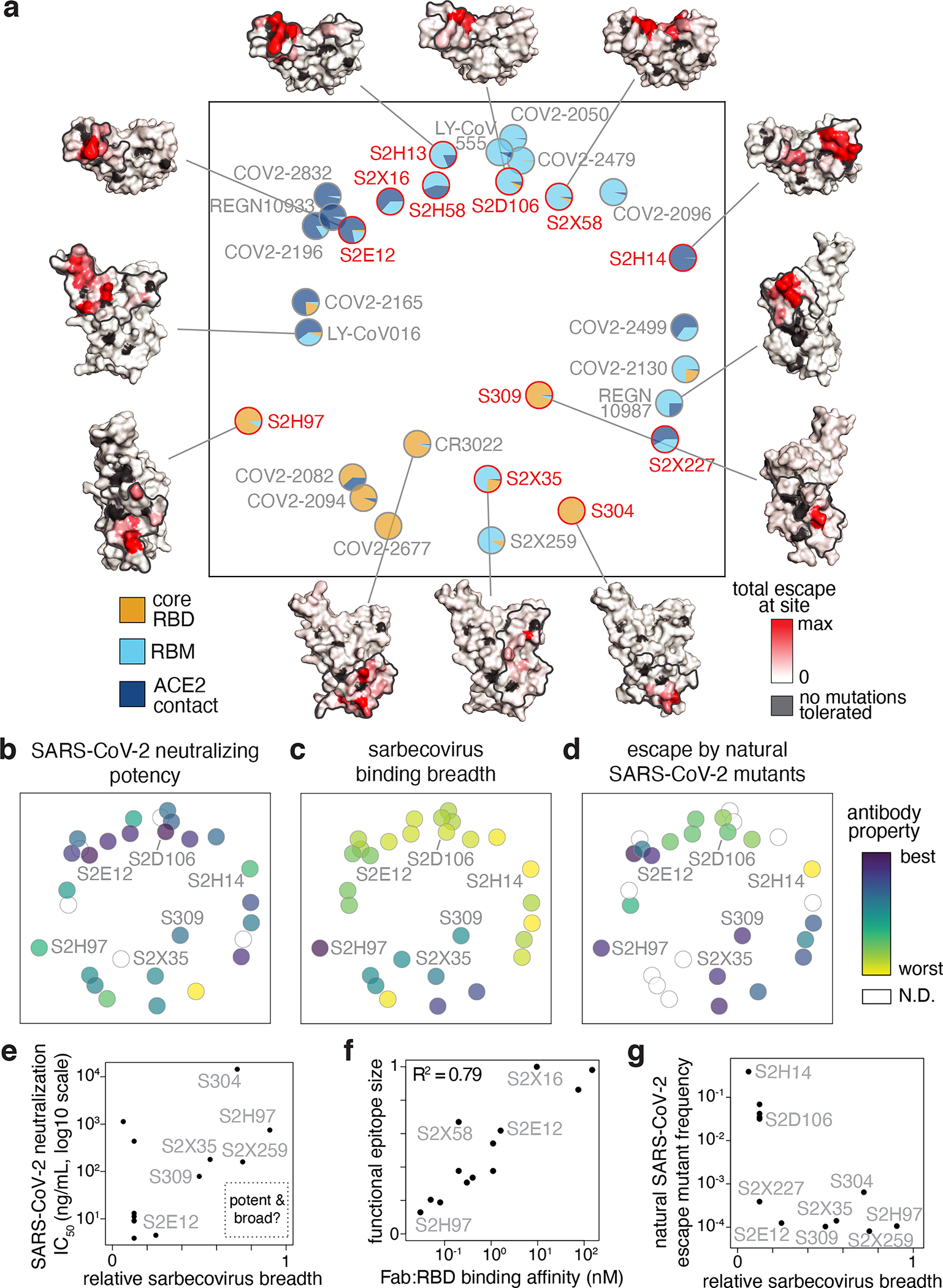
An ideal therapeutic anti-SARS-CoV-2 antibody would resist viral escape1-3, have activity against diverse sarbecoviruses4-7, and be highly protective through viral neutralization8-11 and effector functions12,13. Understanding how these properties relate to each other and vary across epitopes would aid the development of therapeutic antibodies and guide vaccine design. Here we comprehensively characterize escape, breadth and potency across a panel of SARS-CoV-2 antibodies targeting the receptor-binding domain (RBD). Despite a trade-off between in vitro neutralization potency and breadth of sarbecovirus binding, we identify neutralizing antibodies with exceptional sarbecovirus breadth and a corresponding resistance to SARS-CoV-2 escape. One of these antibodies, S2H97, binds with high affinity across all sarbecovirus clades to a cryptic epitope and prophylactically protects hamsters from viral challenge. Antibodies that target the angiotensin-converting enzyme 2 (ACE2) receptor-binding motif (RBM) typically have poor breadth and are readily escaped by mutations despite high neutralization potency. Nevertheless, we also characterize a potent RBM antibody (S2E128) with breadth across sarbecoviruses related to SARS-CoV-2 and a high barrier to viral escape. These data highlight principles underlying variation in escape, breadth and potency among antibodies that target the RBD, and identify epitopes and features to prioritize for therapeutic development against the current and potential future pandemics.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
DECLARATION OF INTERESTS
NC, FZ, DP, MB, PH, RM, JAW, ADM, LER, JZ, MMR, HK, JD, HT, JB, CSF, MPH, JDI, GL, MA, NS, KC, SJ, MM, ED, EC, CHD, LP, FB, AT, FAL, MSP, CMH, HWV, DC and GS are or were employees of Vir Biotechnology and may hold shares in Vir Biotechnology. DC is currently listed as an inventor on multiple patent applications, which disclose the subject matter described in this manuscript. After the submission of the initial version of this study, JDB began consulting for Moderna on viral evolution and epidemiology. JDB has the potential to receive a share of IP revenue as an inventor on a Fred Hutchinson Cancer Research Center-optioned technology/patent (application WO2020006494) related to deep mutational scanning of viral proteins. HWV is a founder of PierianDx and Casma Therapeutics. Neither company provided funding for this work nor is performing related work. JCN, TIC, and DV are consultants for Vir Biotechnology Inc. The Veesler laboratory has received a sponsored research agreement from Vir Biotechnology Inc. JDC is a current member of the Scientific Advisory Boards of OpenEye Scientific Software, Interline Therapeutics, and Redesign Science. The Chodera laboratory receives or has received funding from the National Institute of Health, the National Science Foundation, the Parker Institute for Cancer Immunotherapy, Relay Therapeutics, Entasis Therapeutics, Silicon Therapeutics, EMD Serono (Merck KGaA), AstraZeneca, Vir Biotechnology, XtalPi, Interline Therapeutics, and the Molecular Sciences Software Institute, the Starr Cancer Consortium, the Open Force Field Consortium, Cycle for Survival, a Louis V. Gerstner Young Investigator Award, and the Sloan Kettering Institute. A complete funding history for the Chodera lab can be found at
Figures
Update of
-
Antibodies to the SARS-CoV-2 receptor-binding domain that maximize breadth and resistance to viral escape.bioRxiv [Preprint]. 2021 Apr 8:2021.04.06.438709. doi: 10.1101/2021.04.06.438709. bioRxiv. 2021. Update in: Nature. 2021 Sep;597(7874):97-102. doi: 10.1038/s41586-021-03807-6. PMID: 33851154 Free PMC article. Updated. Preprint.
Comment in
-
Broadly neutralizing anti-coronavirus antibodies.Nat Rev Drug Discov. 2021 Sep;20(9):665. doi: 10.1038/d41573-021-00128-1. Nat Rev Drug Discov. 2021. PMID: 34316026 No abstract available.
References
-
- Pinto D et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 583, 290–295 (2020). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM120553/GM/NIGMS NIH HHS/United States
- DP1 AI158186/AI/NIAID NIH HHS/United States
- P30 CA015704/CA/NCI NIH HHS/United States
- P30 GM124169/GM/NIGMS NIH HHS/United States
- S10 OD023476/OD/NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P30 GM133894/GM/NIGMS NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- R01 AI127893/AI/NIAID NIH HHS/United States
- T32 AI083203/AI/NIAID NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 GM121505/GM/NIGMS NIH HHS/United States
- 209407/Z/17/Z/WT_/Wellcome Trust/United Kingdom
- P30 GM124169-01/NH/NIH HHS/United States
- S10 OD028685/OD/NIH HHS/United States
- R01 GM132386/GM/NIGMS NIH HHS/United States
- HHSN272201700059C/AI/NIAID NIH HHS/United States
- R01 AI141707/AI/NIAID NIH HHS/United States
- T32 GM136534/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
