

cGAS-like receptors sense RNA and control 3'2'-cGAMP signalling in Drosophila
- PMID: 34261127
- PMCID: PMC8410604
- DOI: 10.1038/s41586-021-03743-5
cGAS-like receptors sense RNA and control 3'2'-cGAMP signalling in Drosophila
Abstract
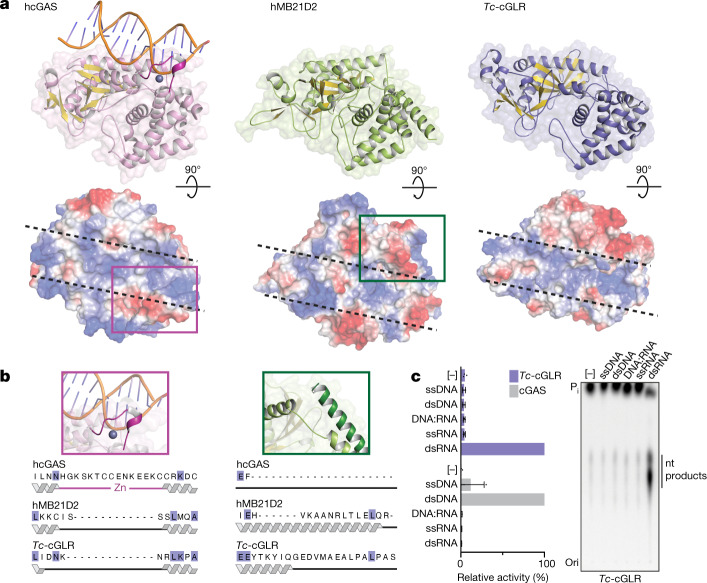
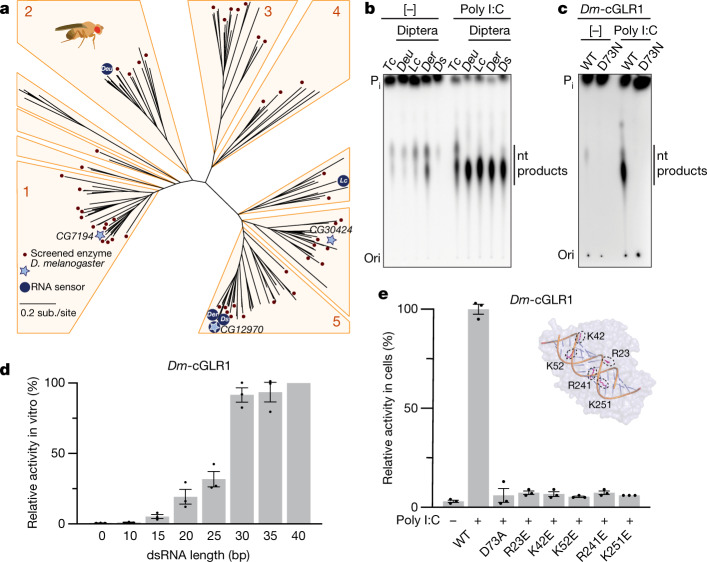
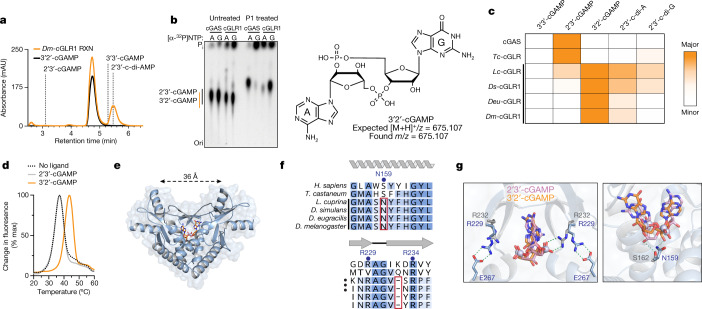
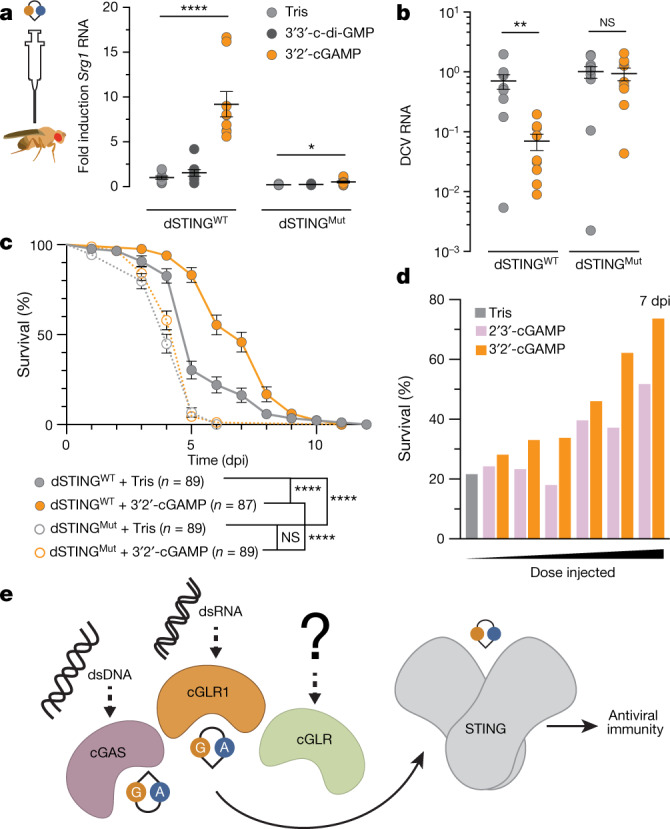
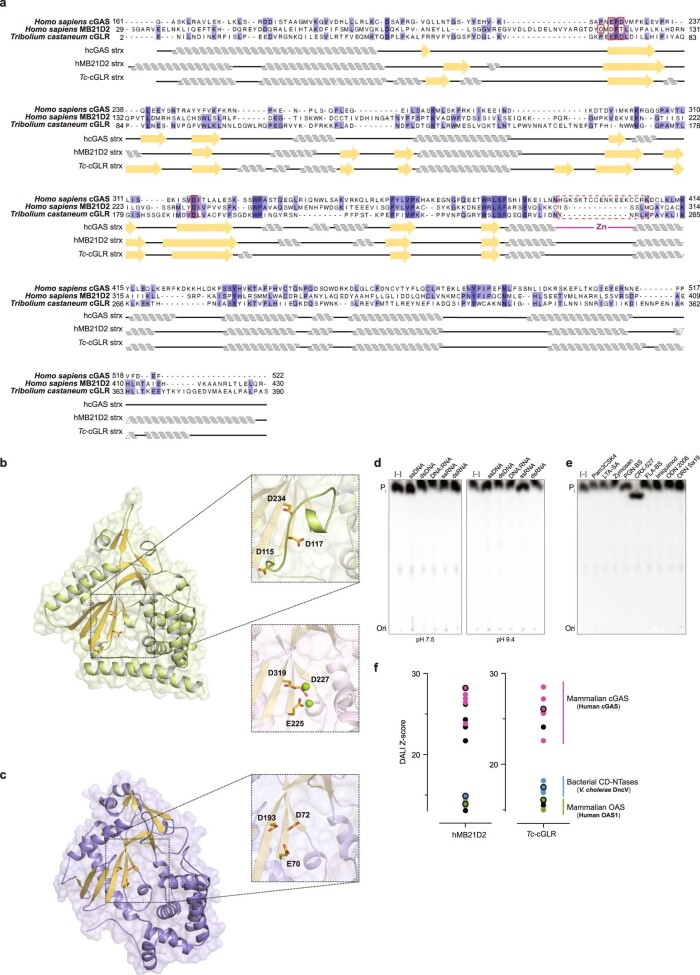
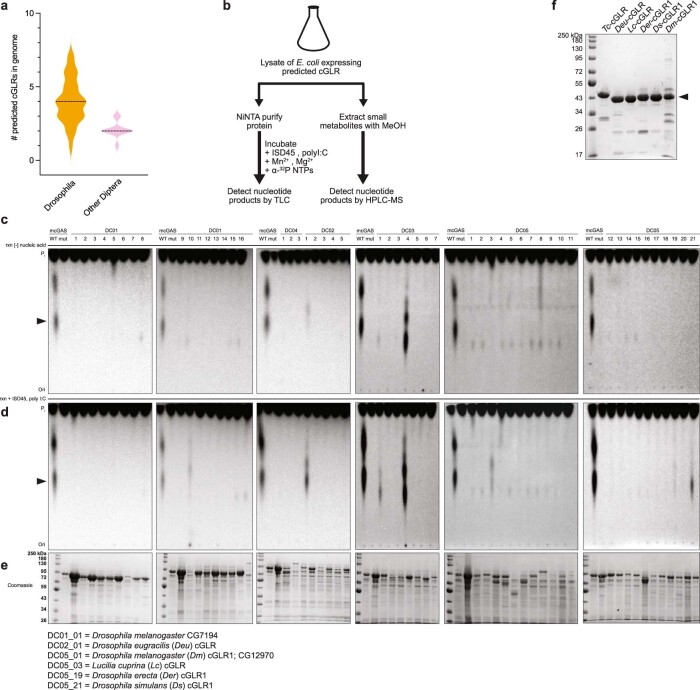
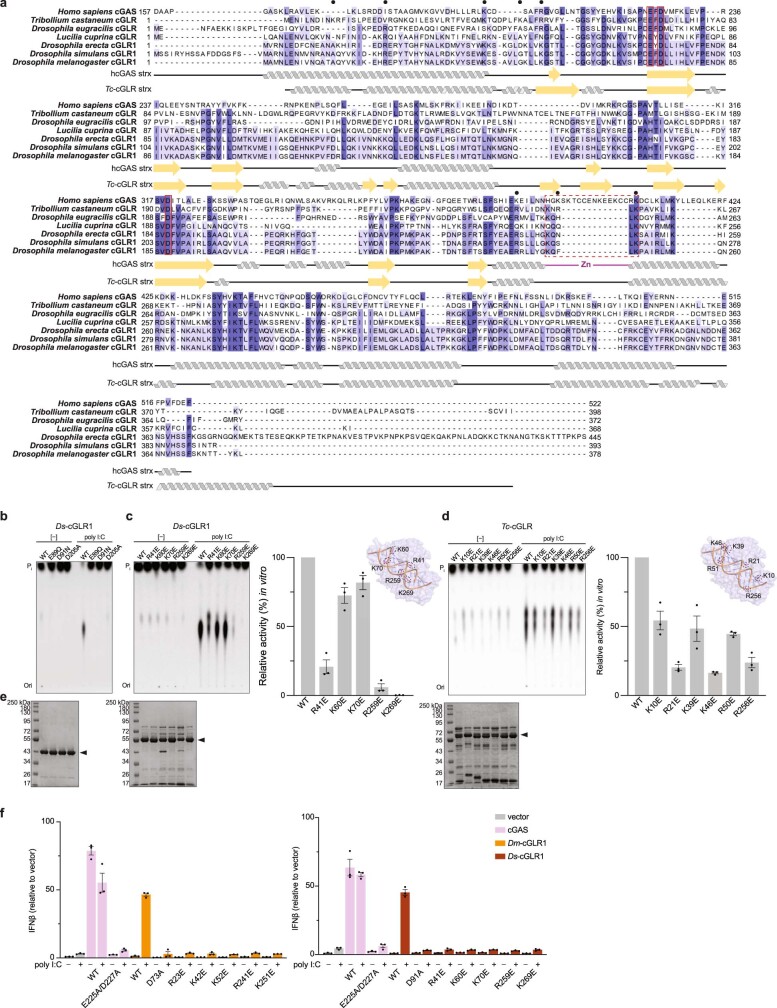
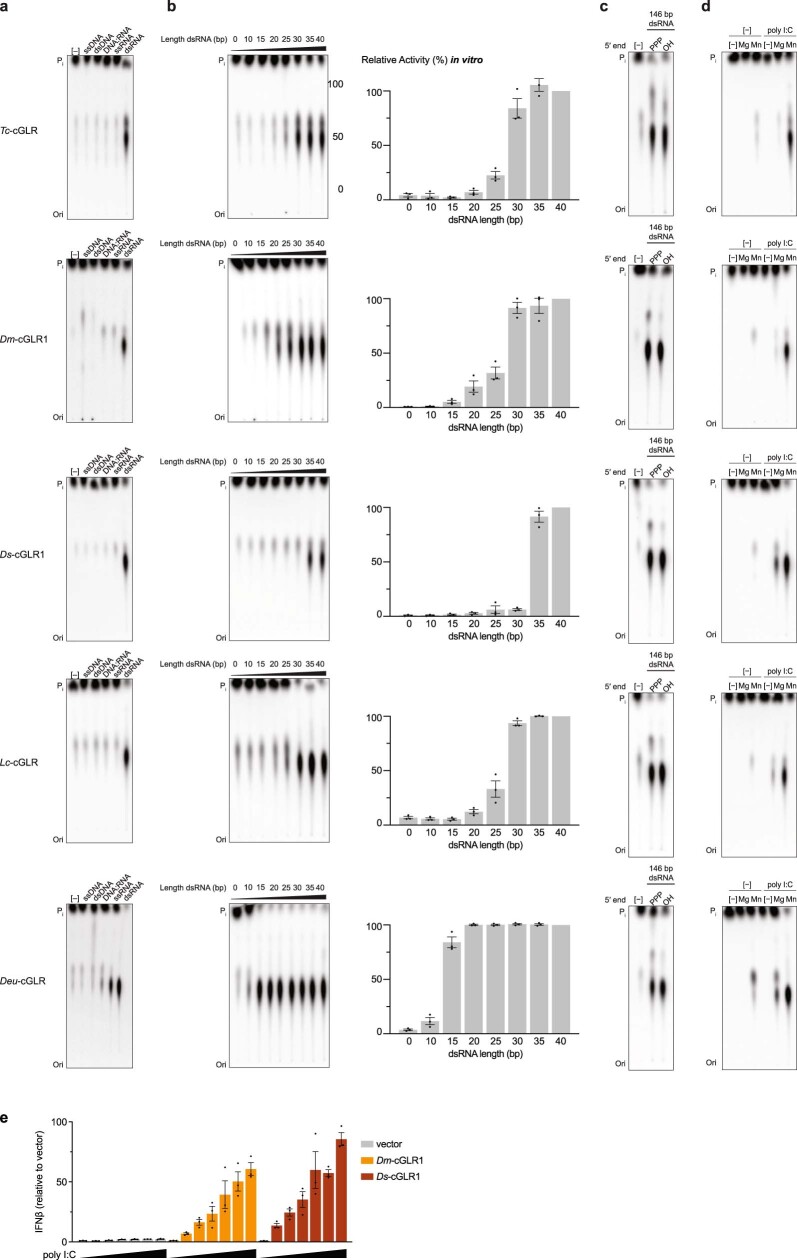
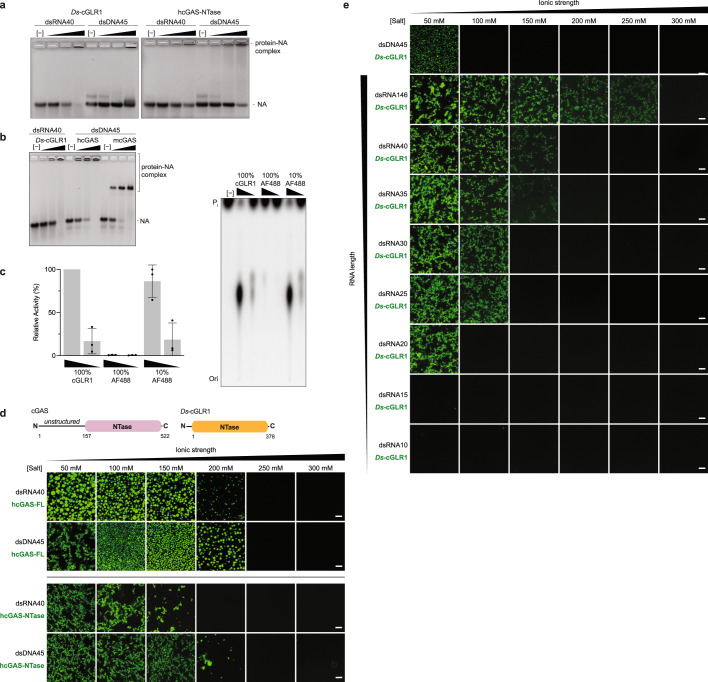
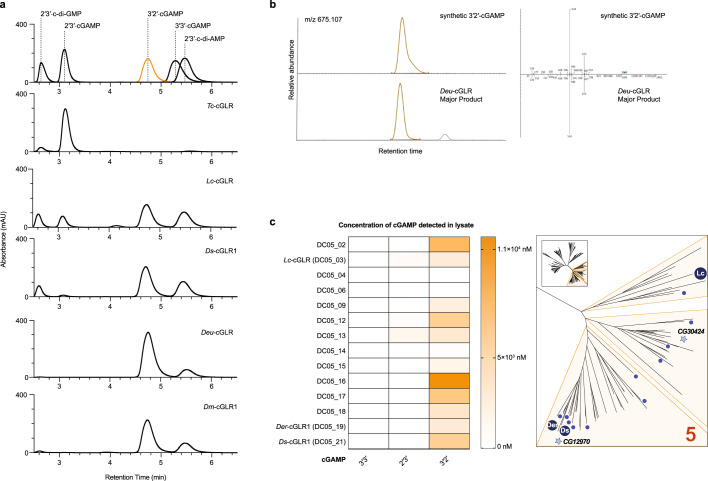
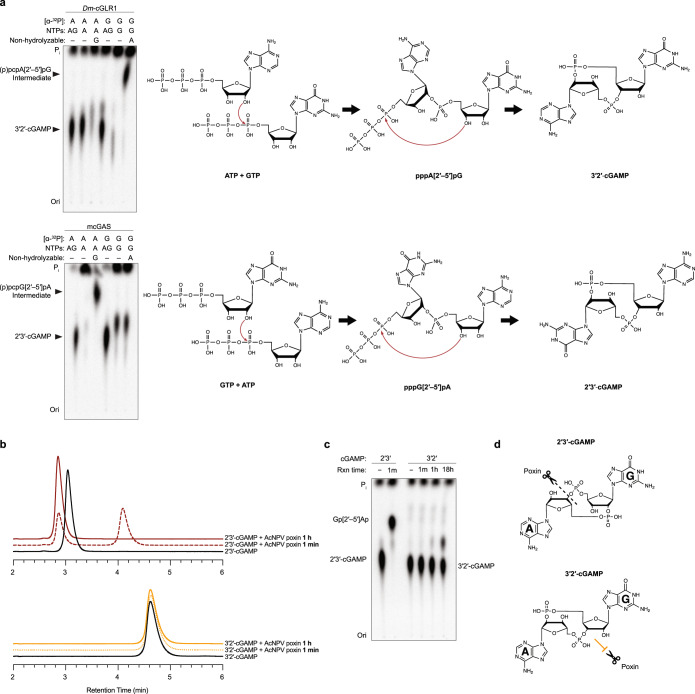
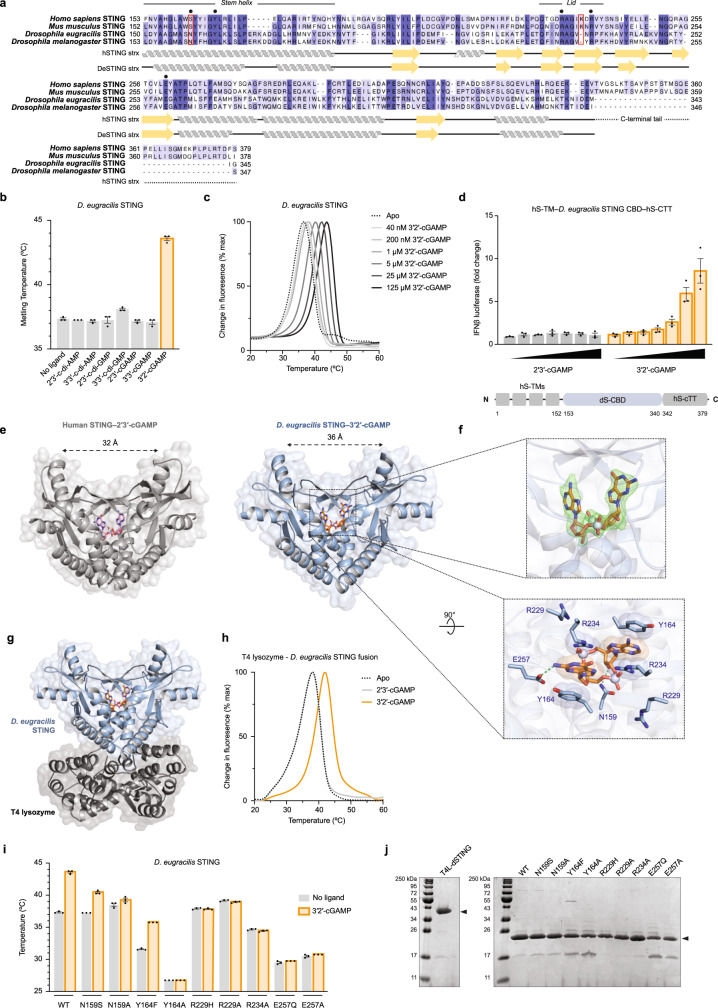
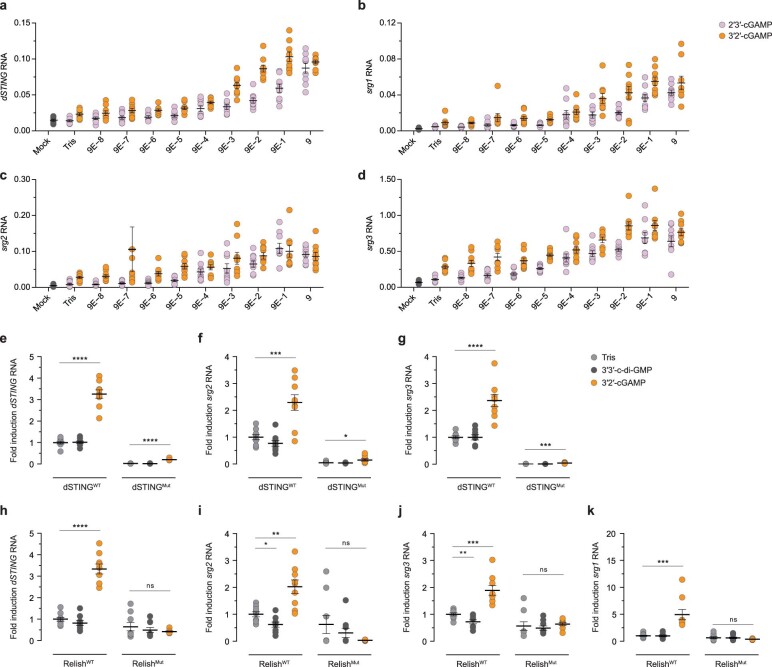
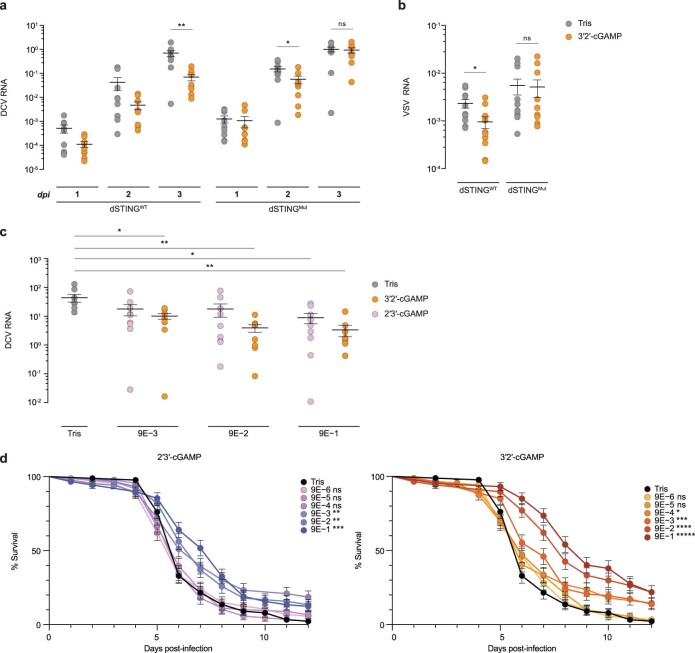
Cyclic GMP-AMP synthase (cGAS) is a cytosolic DNA sensor that produces the second messenger cG[2'-5']pA[3'-5']p (2'3'-cGAMP) and controls activation of innate immunity in mammalian cells1-5. Animal genomes typically encode multiple proteins with predicted homology to cGAS6-10, but the function of these uncharacterized enzymes is unknown. Here we show that cGAS-like receptors (cGLRs) are innate immune sensors that are capable of recognizing divergent molecular patterns and catalysing synthesis of distinct nucleotide second messenger signals. Crystal structures of human and insect cGLRs reveal a nucleotidyltransferase signalling core shared with cGAS and a diversified primary ligand-binding surface modified with notable insertions and deletions. We demonstrate that surface remodelling of cGLRs enables altered ligand specificity and used a forward biochemical screen to identify cGLR1 as a double-stranded RNA sensor in the model organism Drosophila melanogaster. We show that RNA recognition activates Drosophila cGLR1 to synthesize the novel product cG[3'-5']pA[2'-5']p (3'2'-cGAMP). A crystal structure of Drosophila stimulator of interferon genes (dSTING) in complex with 3'2'-cGAMP explains selective isomer recognition, and 3'2'-cGAMP induces an enhanced antiviral state in vivo that protects from viral infection. Similar to radiation of Toll-like receptors in pathogen immunity, our results establish cGLRs as a diverse family of metazoan pattern recognition receptors.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
cGAS-like receptors put a sting into the evolution of immune defence.Nature. 2021 Sep;597(7874):34-35. doi: 10.1038/d41586-021-02119-z. Nature. 2021. PMID: 34373639 No abstract available.
-
cGAS-like receptors: new RNA sensors in Drosophila.Signal Transduct Target Ther. 2021 Oct 25;6(1):370. doi: 10.1038/s41392-021-00785-z. Signal Transduct Target Ther. 2021. PMID: 34697297 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
