

Interactions and Feedbacks in E-Cadherin Transcriptional Regulation
- PMID: 34262912
- PMCID: PMC8273600
- DOI: 10.3389/fcell.2021.701175
Interactions and Feedbacks in E-Cadherin Transcriptional Regulation
Abstract
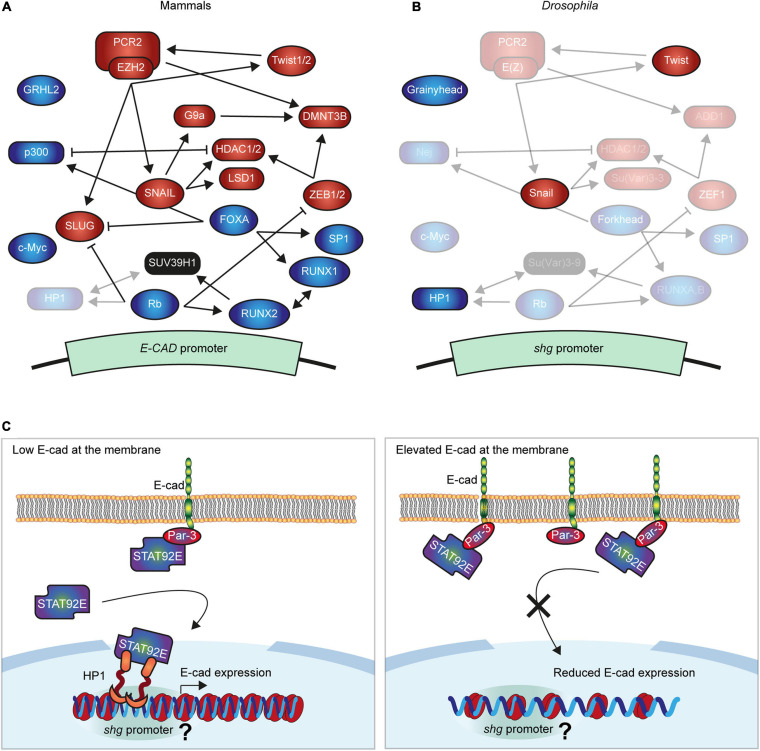
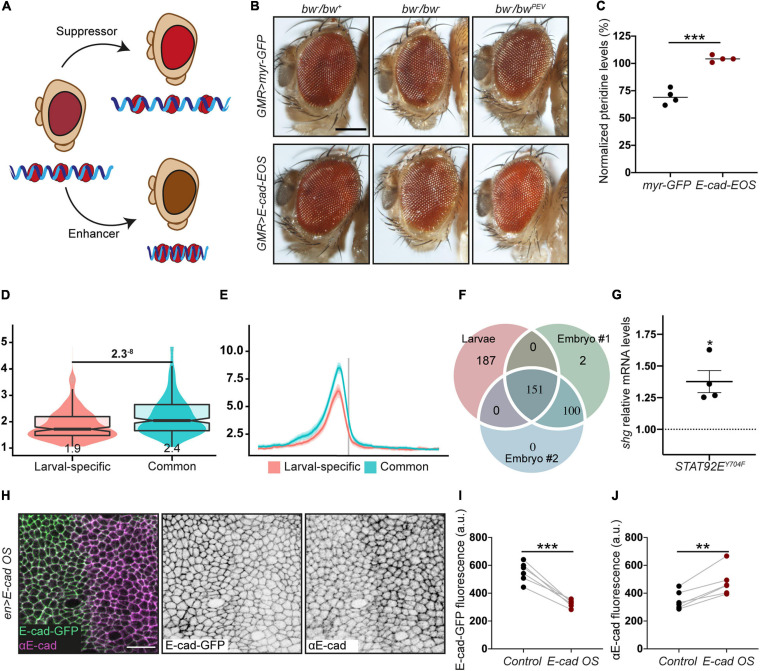
Epithelial tissues rely on the adhesion between participating cells to retain their integrity. The transmembrane protein E-cadherin is the major protein that mediates homophilic adhesion between neighbouring cells and is, therefore, one of the critical components for epithelial integrity. E-cadherin downregulation has been described extensively as a prerequisite for epithelial-to-mesenchymal transition and is a hallmark in many types of cancer. Due to this clinical importance, research has been mostly focused on understanding the mechanisms leading to transcriptional repression of this adhesion molecule. However, in recent years it has become apparent that re-expression of E-cadherin is a major step in the progression of many cancers during metastasis. Here, we review the currently known molecular mechanisms of E-cadherin transcriptional activation and inhibition and highlight complex interactions between individual mechanisms. We then propose an additional mechanism, whereby the competition between adhesion complexes and heterochromatin protein-1 for binding to STAT92E fine-tunes the levels of E-cadherin expression in Drosophila but also regulates other genes promoting epithelial robustness. We base our hypothesis on both existing literature and our experimental evidence and suggest that such feedback between the cell surface and the nucleus presents a powerful paradigm for epithelial resilience.
Keywords: HP1; JAK/STAT; PAR-3; adhesion; heterochromatin.
Copyright © 2021 Ramirez Moreno, Stempor and Bulgakova.
Conflict of interest statement
PAS was a shareholder and the managing director of the company SmartImmune Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Prospero homeobox 1 promotes epithelial-mesenchymal transition in colon cancer cells by inhibiting E-cadherin via miR-9.Clin Cancer Res. 2012 Dec 1;18(23):6416-25. doi: 10.1158/1078-0432.CCR-12-0832. Epub 2012 Oct 8. Clin Cancer Res. 2012. PMID: 23045246
-
Posttranslational truncation of E-cadherin and significance for tumour progression.Cells Tissues Organs. 2007;185(1-3):175-9. doi: 10.1159/000101318. Cells Tissues Organs. 2007. PMID: 17587823 Review.
-
Inflammatory mediators drive metastasis and drug resistance in head and neck squamous cell carcinoma.Laryngoscope. 2015 Mar;125 Suppl 3:S1-11. doi: 10.1002/lary.24998. Epub 2015 Feb 3. Laryngoscope. 2015. PMID: 25646683
-
PRRX1 deficiency induces mesenchymal-epithelial transition through PITX2/miR-200-dependent SLUG/CTNNB1 regulation in hepatocellular carcinoma.Cancer Sci. 2021 Jun;112(6):2158-2172. doi: 10.1111/cas.14853. Epub 2021 Apr 8. Cancer Sci. 2021. PMID: 33587761 Free PMC article.
-
Cadherin profiling for therapeutic interventions in Epithelial Mesenchymal Transition (EMT) and tumorigenesis.Exp Cell Res. 2018 Jul 15;368(2):137-146. doi: 10.1016/j.yexcr.2018.04.014. Epub 2018 Apr 16. Exp Cell Res. 2018. PMID: 29674112 Review.
Cited by
-
Reversible Photoregulation of Cell-Cell Adhesions With Opto-E-cadherin.Bio Protoc. 2024 May 20;14(10):e4995. doi: 10.21769/BioProtoc.4995. eCollection 2024 May 20. Bio Protoc. 2024. PMID: 38798983 Free PMC article.
-
E-cadherin endocytosis promotes non-canonical EGFR:STAT signalling to induce cell death and inhibit heterochromatinisation.PLoS Genet. 2025 Jul 21;21(7):e1011781. doi: 10.1371/journal.pgen.1011781. eCollection 2025 Jul. PLoS Genet. 2025. PMID: 40690511 Free PMC article.
-
HOXD1 regulates neural crest cells differentiation and polycerate development in sheep.Sci Rep. 2025 Apr 16;15(1):13140. doi: 10.1038/s41598-025-97865-9. Sci Rep. 2025. PMID: 40240791 Free PMC article.
-
E-cadherin acts as a positive regulator of the JAK-STAT signaling pathway during Drosophila oogenesis.Front Cell Dev Biol. 2022 Aug 23;10:886312. doi: 10.3389/fcell.2022.886312. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36120588 Free PMC article.
-
Identification of a core EMT signature that separates basal-like breast cancers into partial- and post-EMT subtypes.Front Oncol. 2023 Dec 4;13:1249895. doi: 10.3389/fonc.2023.1249895. eCollection 2023. Front Oncol. 2023. PMID: 38111531 Free PMC article.
References
-
- Arima Y., Inoue Y., Shibata T., Hayashi H., Nagano O., Saya H., et al. (2008). Rb depletion results in deregulation of E-cadherin and induction of cellular phenotypic changes that are characteristic of the epithelial-to-mesenchymal transition. Cancer Res. 68 5104–5112. 10.1158/0008-5472.can-07-5680 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials