

Multifaceted roles of centrosomes in development, health, and disease
- PMID: 34264337
- PMCID: PMC8648388
- DOI: 10.1093/jmcb/mjab041
Multifaceted roles of centrosomes in development, health, and disease
Abstract
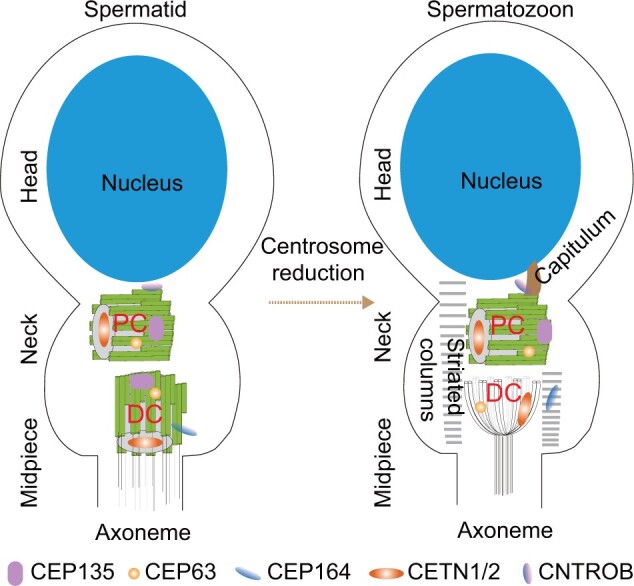
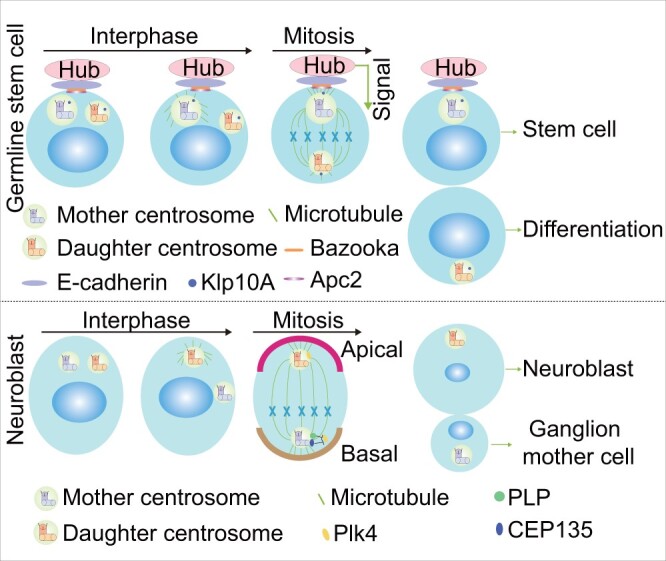
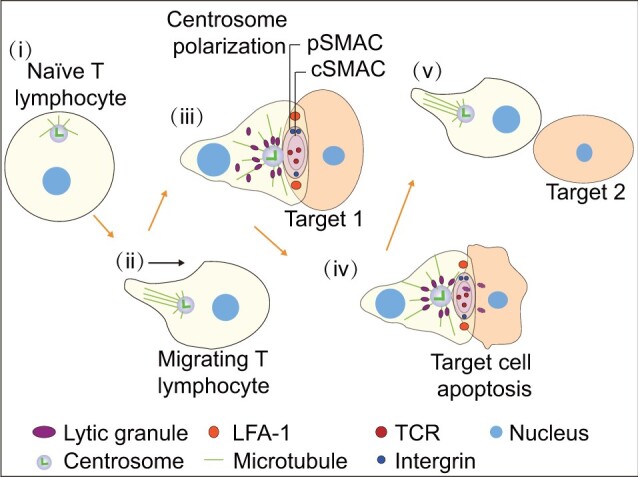
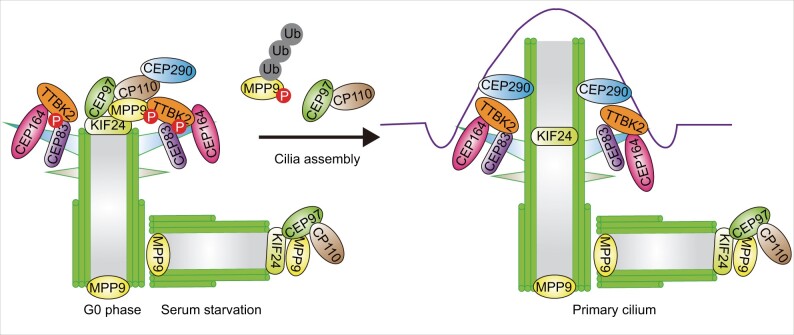
The centrosome is a membrane-less organelle consisting of a pair of barrel-shaped centrioles and pericentriolar material and functions as the major microtubule-organizing center and signaling hub in animal cells. The past decades have witnessed the functional complexity and importance of centrosomes in various cellular processes such as cell shaping, division, and migration. In addition, centrosome abnormalities are linked to a wide range of human diseases and pathological states, such as cancer, reproductive disorder, brain disease, and ciliopathies. Herein, we discuss various functions of centrosomes in development and health, with an emphasis on their roles in germ cells, stem cells, and immune responses. We also discuss how centrosome dysfunctions are involved in diseases. A better understanding of the mechanisms regulating centrosome functions may lead the way to potential therapeutic targeting of this organelle in disease treatment.
Keywords: centrosome; ciliopathy; germ cell; immunity; stem cell.
© The Author(s) (2021). Published by Oxford University Press on behalf of Journal of Molecular Cell Biology, CEMCS, CAS.
Figures
References
-
- Ansley S.J., Badano J.L., Blacque O.E., et al. (2003). Basal body dysfunction is a likely cause of pleiotropic Bardet‒Biedl syndrome. Nature 425, 628–633. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
