

A neural code for egocentric spatial maps in the human medial temporal lobe
- PMID: 34265253
- PMCID: PMC8518022
- DOI: 10.1016/j.neuron.2021.06.019
A neural code for egocentric spatial maps in the human medial temporal lobe
Abstract
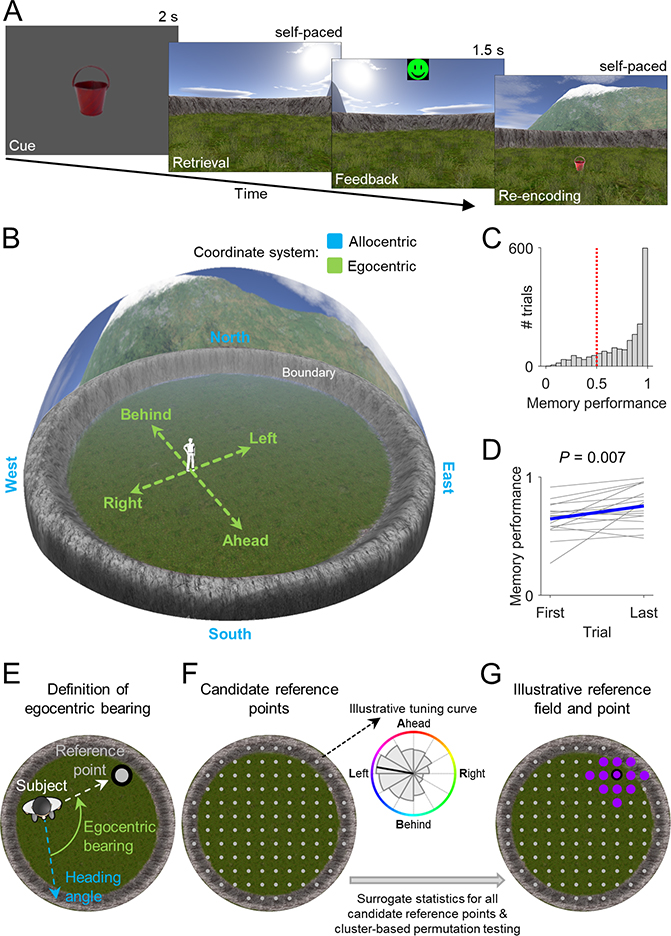
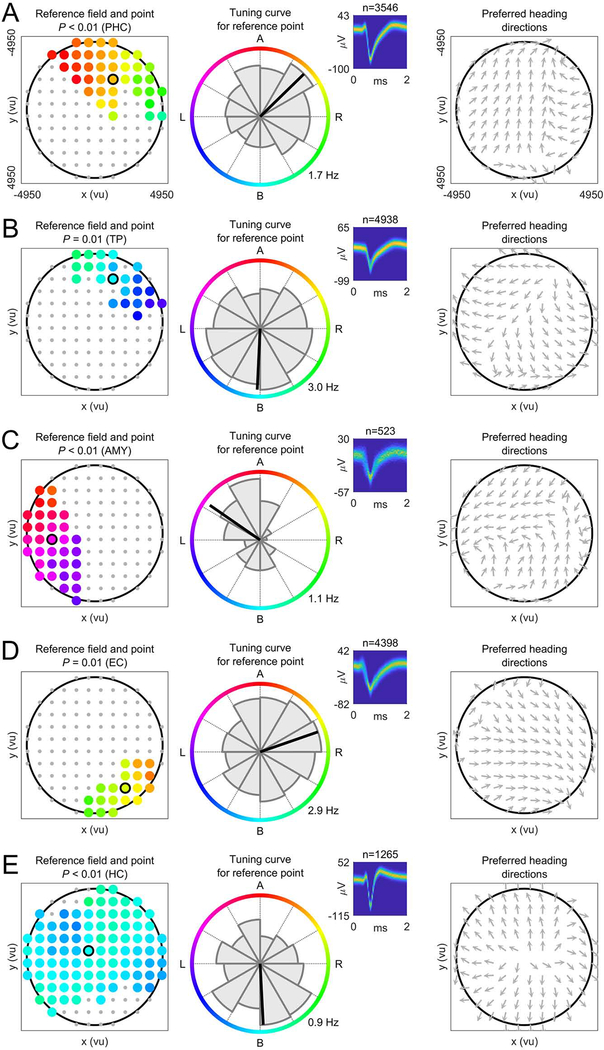
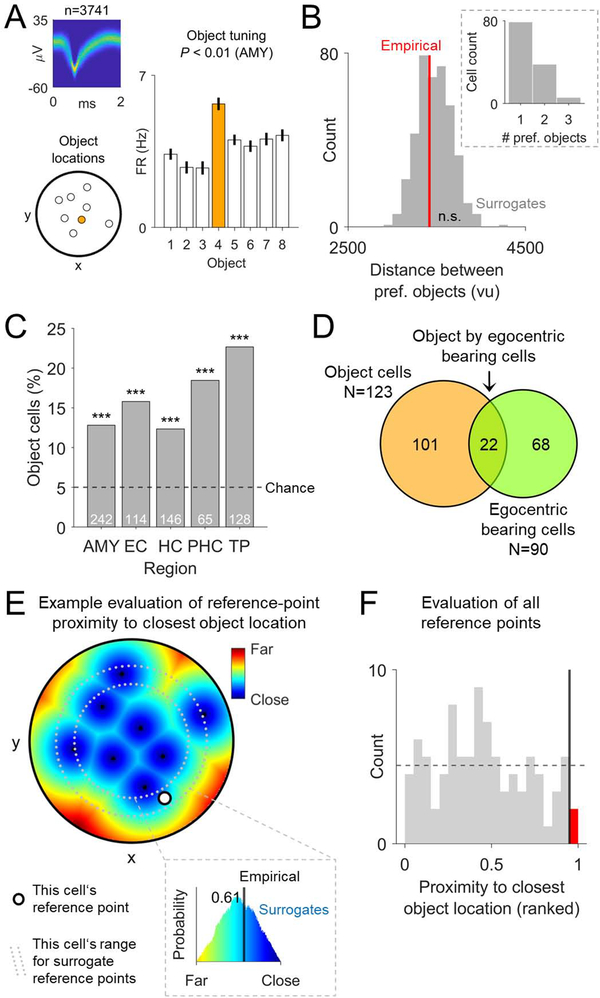
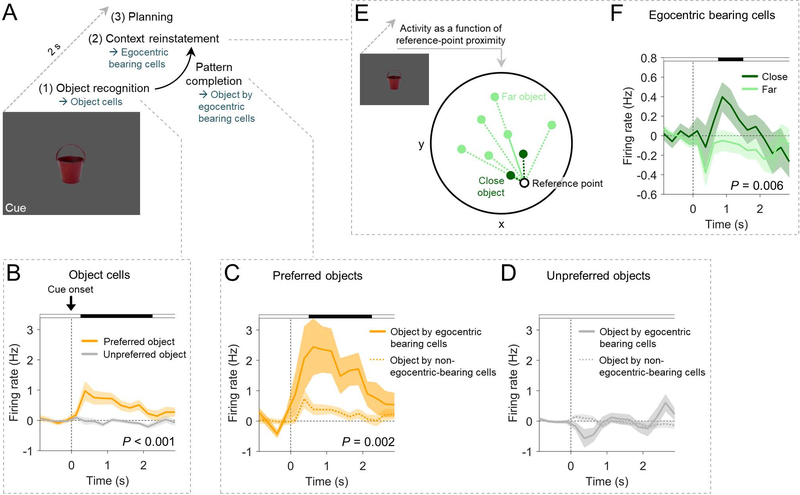
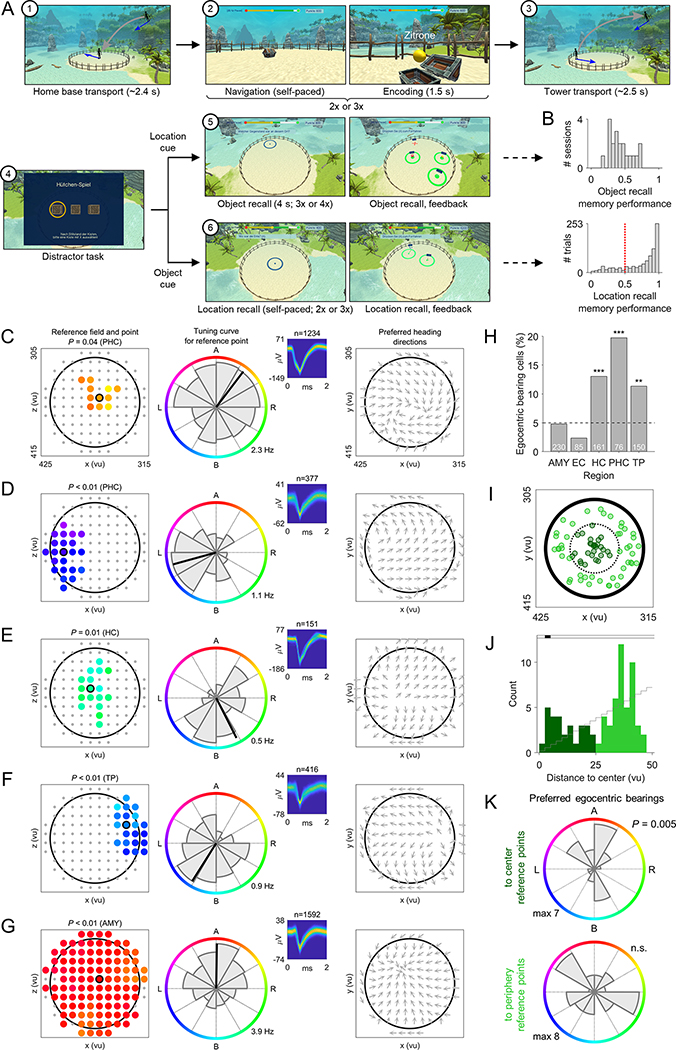
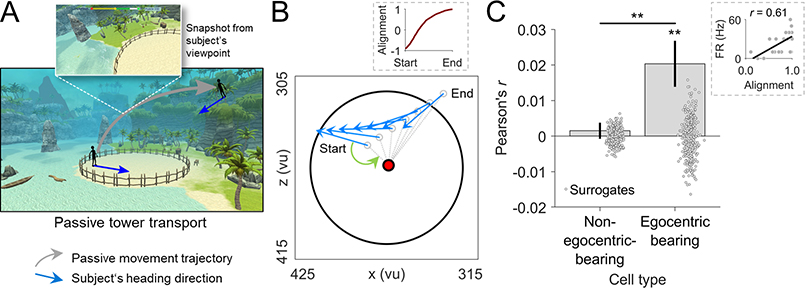
Spatial navigation and memory rely on neural systems that encode places, distances, and directions in relation to the external world or relative to the navigating organism. Place, grid, and head-direction cells form key units of world-referenced, allocentric cognitive maps, but the neural basis of self-centered, egocentric representations remains poorly understood. Here, we used human single-neuron recordings during virtual spatial navigation tasks to identify neurons providing a neural code for egocentric spatial maps in the human brain. Consistent with previous observations in rodents, these neurons represented egocentric bearings toward reference points positioned throughout the environment. Egocentric bearing cells were abundant in the parahippocampal cortex and supported vectorial representations of egocentric space by also encoding distances toward reference points. Beyond navigation, the observed neurons showed activity increases during spatial and episodic memory recall, suggesting that egocentric bearing cells are not only relevant for navigation but also play a role in human memory.
Keywords: allocentric; egocentric; electrophysiology; hippocampus; human single-neuron recording; memory; navigation; parahippocampal cortex; sense of direction.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


References
-
- Aguirre GK, Detre JA, Alsop DC, and D’Esposito M (1996). The Parahippocampus Subserves Topographical Learning in Man. Cereb. Cortex 6, 823–829. - PubMed
-
- Berens P (2009). CircStat: A MATLAB Toolbox for Circular Statistics. J. Stat. Softw. 31, 1–21.
-
- Boccara CN, Nardin M, Stella F, O’Neill J, and Csicsvari J (2019). The entorhinal cognitive map is attracted to goals. Science 363, 1443–1447. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
