

Knock-in models related to Alzheimer's disease: synaptic transmission, plaques and the role of microglia
- PMID: 34266459
- PMCID: PMC8281661
- DOI: 10.1186/s13024-021-00457-0
Knock-in models related to Alzheimer's disease: synaptic transmission, plaques and the role of microglia
Abstract
Background: Microglia are active modulators of Alzheimer's disease but their role in relation to amyloid plaques and synaptic changes due to rising amyloid beta is unclear. We add novel findings concerning these relationships and investigate which of our previously reported results from transgenic mice can be validated in knock-in mice, in which overexpression and other artefacts of transgenic technology are avoided.
Methods: AppNL-F and AppNL-G-F knock-in mice expressing humanised amyloid beta with mutations in App that cause familial Alzheimer's disease were compared to wild type mice throughout life. In vitro approaches were used to understand microglial alterations at the genetic and protein levels and synaptic function and plasticity in CA1 hippocampal neurones, each in relationship to both age and stage of amyloid beta pathology. The contribution of microglia to neuronal function was further investigated by ablating microglia with CSF1R inhibitor PLX5622.
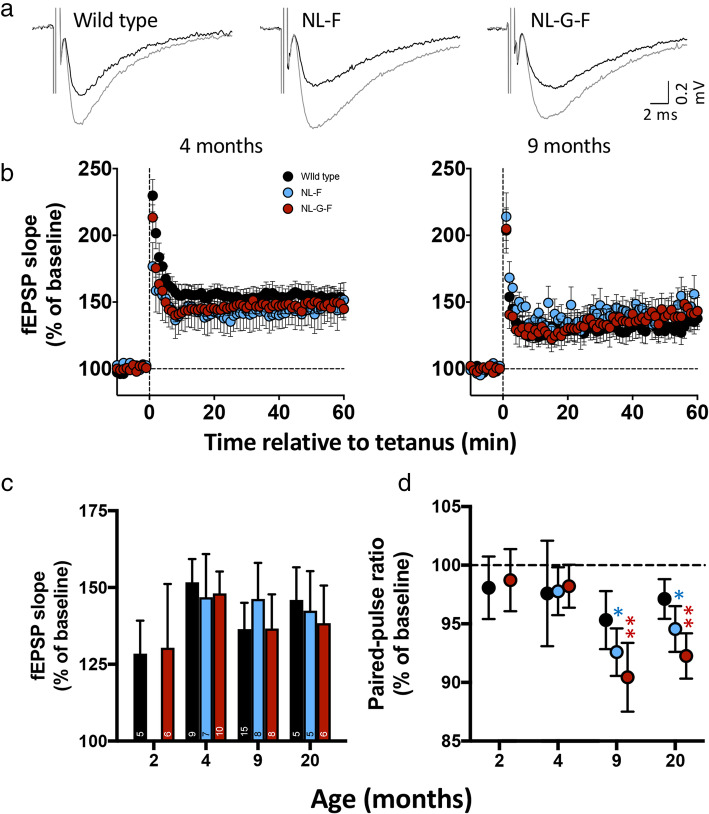
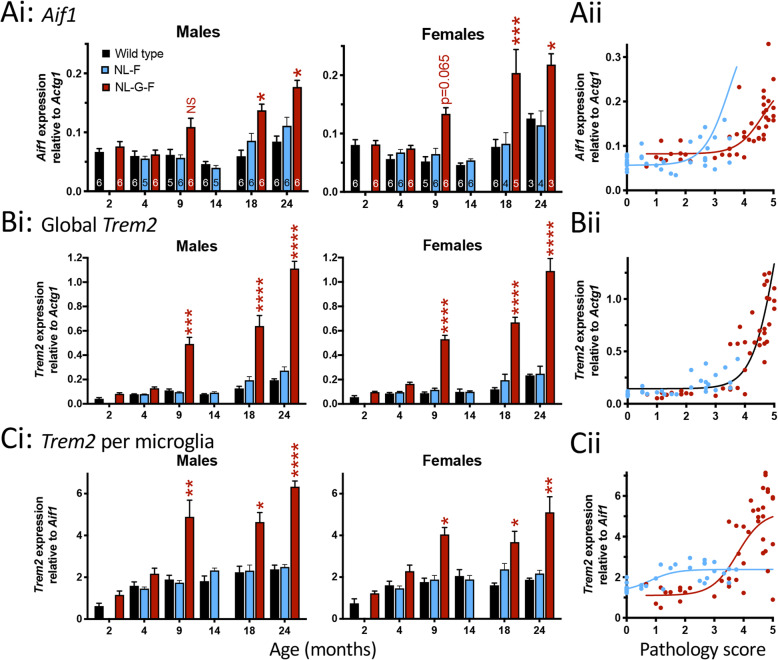
Results: Both App knock-in lines showed increased glutamate release probability prior to detection of plaques. Consistent with results in transgenic mice, this persisted throughout life in AppNL-F mice but was not evident in AppNL-G-F with sparse plaques. Unlike transgenic mice, loss of spontaneous excitatory activity only occurred at the latest stages, while no change could be detected in spontaneous inhibitory synaptic transmission or magnitude of long-term potentiation. Also, in contrast to transgenic mice, the microglial response in both App knock-in lines was delayed until a moderate plaque load developed. Surviving PLX5266-depleted microglia tended to be CD68-positive. Partial microglial ablation led to aged but not young wild type animals mimicking the increased glutamate release probability in App knock-ins and exacerbated the App knock-in phenotype. Complete ablation was less effective in altering synaptic function, while neither treatment altered plaque load.
Conclusions: Increased glutamate release probability is similar across knock-in and transgenic mouse models of Alzheimer's disease, likely reflecting acute physiological effects of soluble amyloid beta. Microglia respond later to increased amyloid beta levels by proliferating and upregulating Cd68 and Trem2. Partial depletion of microglia suggests that, in wild type mice, alteration of surviving phagocytic microglia, rather than microglial loss, drives age-dependent effects on glutamate release that become exacerbated in Alzheimer's disease.
Keywords: Ageing; Alzheimer’s disease; Amyloid beta; Gene expression; Microglia; Neurodegeneration; Plaques; Synaptic plasticity; Synaptic transmission; TREM2.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
