

The DME demethylase regulates sporophyte gene expression, cell proliferation, differentiation, and meristem resurrection
- PMID: 34266952
- PMCID: PMC8307533
- DOI: 10.1073/pnas.2026806118
The DME demethylase regulates sporophyte gene expression, cell proliferation, differentiation, and meristem resurrection
Erratum in
-
Correction for Kim et al., The DME demethylase regulates sporophyte gene expression, cell proliferation, differentiation, and meristem resurrection.Proc Natl Acad Sci U S A. 2023 Aug;120(31):e2311112120. doi: 10.1073/pnas.2311112120. Epub 2023 Jul 24. Proc Natl Acad Sci U S A. 2023. PMID: 37487107 Free PMC article. No abstract available.
Abstract
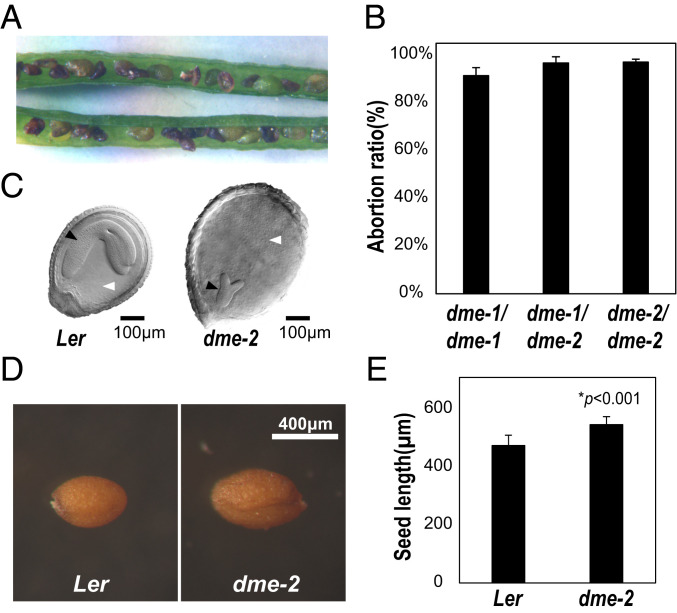
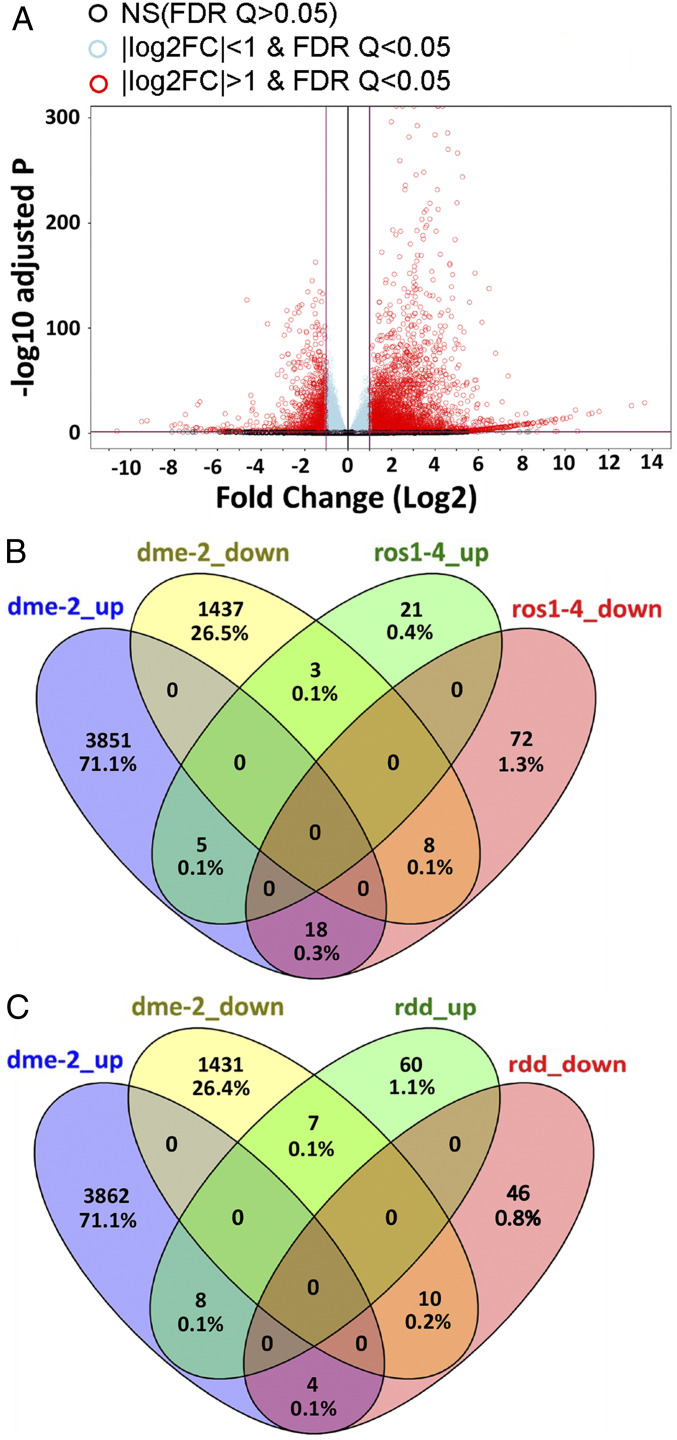
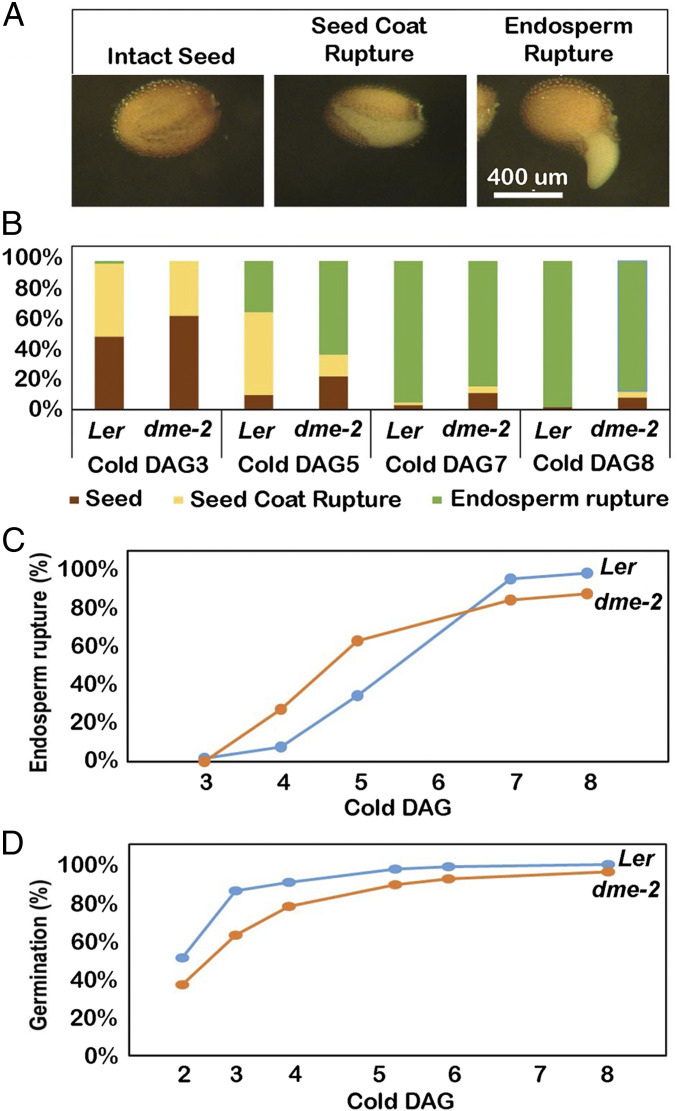
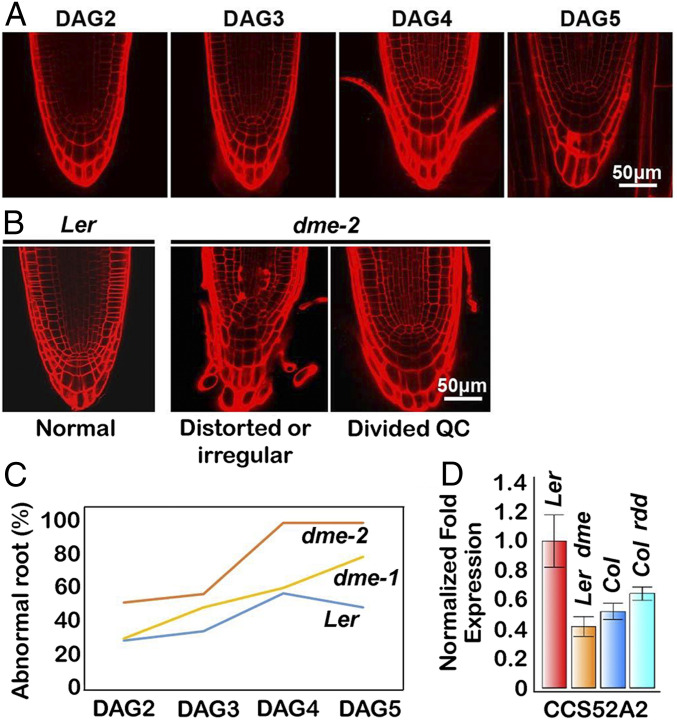
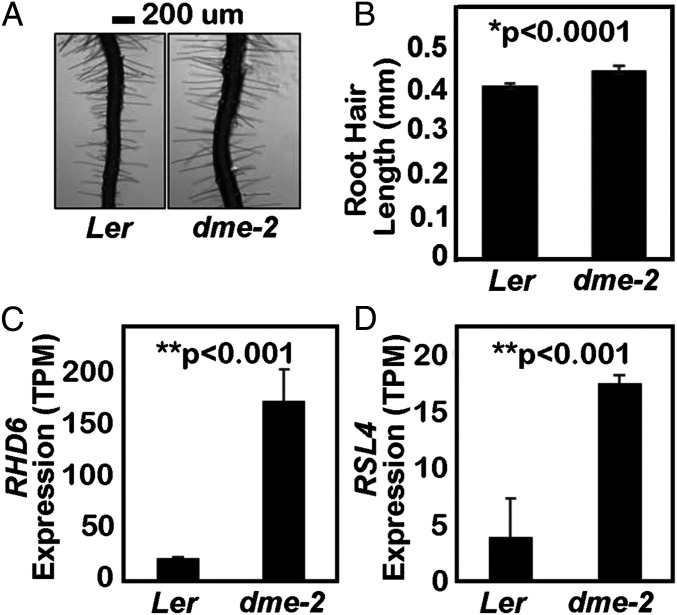
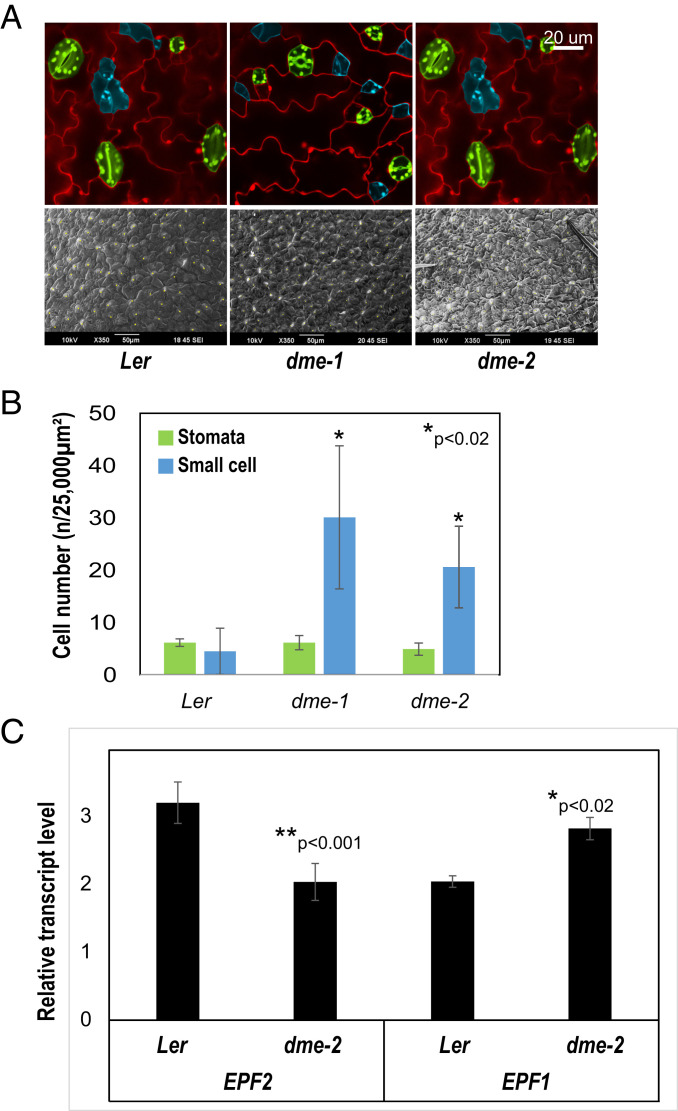
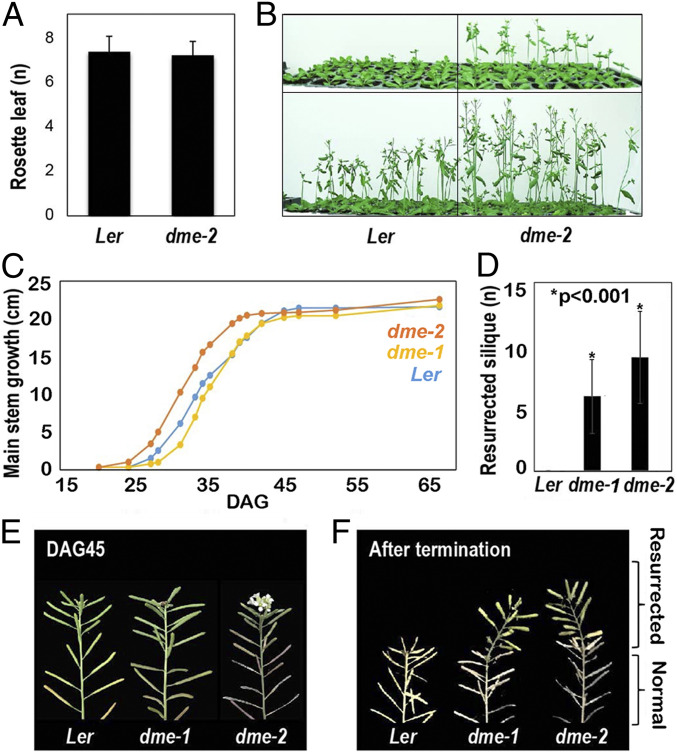
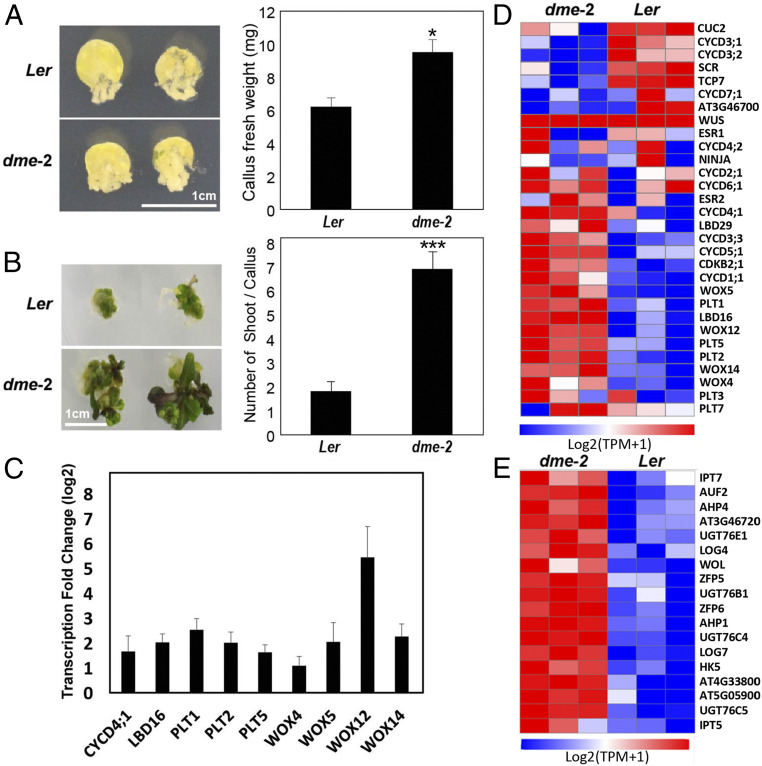
The flowering plant life cycle consists of alternating haploid (gametophyte) and diploid (sporophyte) generations, where the sporophytic generation begins with fertilization of haploid gametes. In Arabidopsis, genome-wide DNA demethylation is required for normal development, catalyzed by the DEMETER (DME) DNA demethylase in the gamete companion cells of male and female gametophytes. In the sporophyte, postembryonic growth and development are largely dependent on the activity of numerous stem cell niches, or meristems. Analyzing Arabidopsis plants homozygous for a loss-of-function dme-2 allele, we show that DME influences many aspects of sporophytic growth and development. dme-2 mutants exhibited delayed seed germination, variable root hair growth, aberrant cellular proliferation and differentiation followed by enhanced de novo shoot formation, dysregulation of root quiescence and stomatal precursor cells, and inflorescence meristem (IM) resurrection. We also show that sporophytic DME activity exerts a profound effect on the transcriptome of developing Arabidopsis plants, including discrete groups of regulatory genes that are misregulated in dme-2 mutant tissues, allowing us to potentially link phenotypes to changes in specific gene expression pathways. These results show that DME plays a key role in sporophytic development and suggest that DME-mediated active DNA demethylation may be involved in the maintenance of stem cell activities during the sporophytic life cycle in Arabidopsis.
Keywords: DNA demethylation; cell proliferation; pluripotency; sprorophytic development.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Gametophytic epigenetic regulators, MEDEA and DEMETER, synergistically suppress ectopic shoot formation in Arabidopsis.Plant Cell Rep. 2024 Feb 11;43(3):68. doi: 10.1007/s00299-024-03159-1. Plant Cell Rep. 2024. PMID: 38341844
-
Control of DEMETER DNA demethylase gene transcription in male and female gamete companion cells in Arabidopsis thaliana.Proc Natl Acad Sci U S A. 2017 Feb 21;114(8):2078-2083. doi: 10.1073/pnas.1620592114. Epub 2017 Jan 27. Proc Natl Acad Sci U S A. 2017. PMID: 28130550 Free PMC article.
-
Function of the DEMETER DNA glycosylase in the Arabidopsis thaliana male gametophyte.Proc Natl Acad Sci U S A. 2011 May 10;108(19):8042-7. doi: 10.1073/pnas.1105117108. Epub 2011 Apr 25. Proc Natl Acad Sci U S A. 2011. PMID: 21518889 Free PMC article.
-
Functions and Regulation of HAM Family Genes in Meristems During Gametophyte and Sporophyte Generations.Plant Cell Environ. 2025 Mar;48(3):2125-2131. doi: 10.1111/pce.15286. Epub 2024 Nov 18. Plant Cell Environ. 2025. PMID: 39558470 Free PMC article. Review.
-
Genome demethylation and imprinting in the endosperm.Curr Opin Plant Biol. 2011 Apr;14(2):162-7. doi: 10.1016/j.pbi.2011.02.006. Epub 2011 Mar 23. Curr Opin Plant Biol. 2011. PMID: 21435940 Free PMC article. Review.
Cited by
-
The pivotal ripening gene SlDML2 participates in regulating disease resistance in tomato.Plant Biotechnol J. 2023 Nov;21(11):2291-2306. doi: 10.1111/pbi.14130. Epub 2023 Jul 19. Plant Biotechnol J. 2023. PMID: 37466912 Free PMC article.
-
Molecular mechanisms and biological functions of active DNA demethylation in plants.Epigenetics Chromatin. 2025 Jul 5;18(1):41. doi: 10.1186/s13072-025-00605-6. Epigenetics Chromatin. 2025. PMID: 40618161 Free PMC article. Review.
-
Effects of vitrectomy combined with internal limiting membrane peeling in patients with diabetic macular edema.World J Clin Cases. 2024 Jul 26;12(21):4491-4498. doi: 10.12998/wjcc.v12.i21.4491. World J Clin Cases. 2024. PMID: 39070848 Free PMC article.
-
CG and CHG Methylation Contribute to the Transcriptional Control of OsPRR37-Output Genes in Rice.Front Plant Sci. 2022 Feb 15;13:839457. doi: 10.3389/fpls.2022.839457. eCollection 2022. Front Plant Sci. 2022. PMID: 35242159 Free PMC article.
-
Epigenetic Regulation During Plant Development and the Capacity for Epigenetic Memory.Annu Rev Plant Biol. 2023 May 22;74:87-109. doi: 10.1146/annurev-arplant-070122-025047. Epub 2023 Feb 28. Annu Rev Plant Biol. 2023. PMID: 36854474 Free PMC article. Review.
References
-
- Hofmeister W. F. B., Currey F., On the Germination, Development, and Fructification of the Higher Cryptogamia, and On the Fructification of the Coniferæ;, The Ray society (Publication for the Ray society by R. Hardwicke, London, 1862).
-
- Choi Y., et al. ., DEMETER, a DNA glycosylase domain protein, is required for endosperm gene imprinting and seed viability in arabidopsis. Cell 110, 33–42 (2002). - PubMed
