

Membranotropic and biological activities of the membrane fusion peptides from SARS-CoV spike glycoprotein: The importance of the complete internal fusion peptide domain
- PMID: 34274319
- PMCID: PMC8280623
- DOI: 10.1016/j.bbamem.2021.183697
Membranotropic and biological activities of the membrane fusion peptides from SARS-CoV spike glycoprotein: The importance of the complete internal fusion peptide domain
Abstract
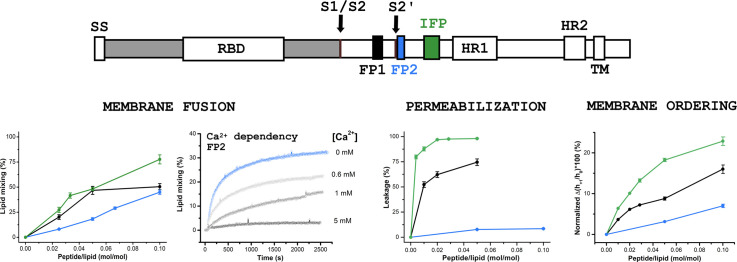
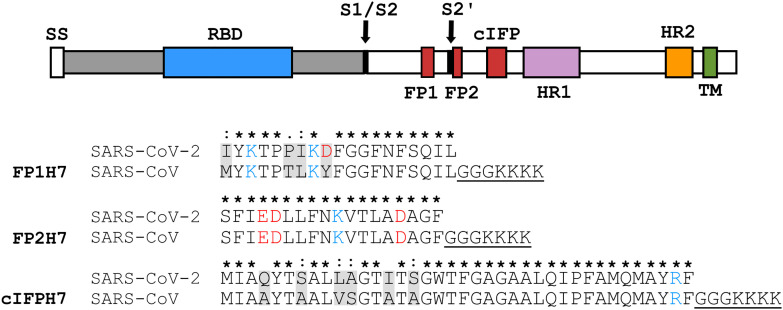
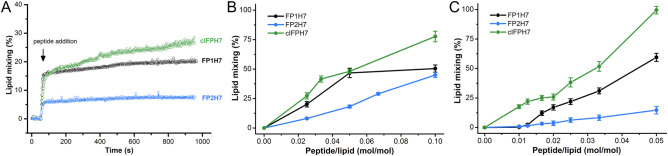
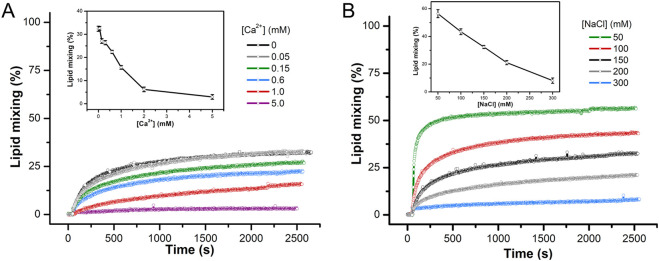
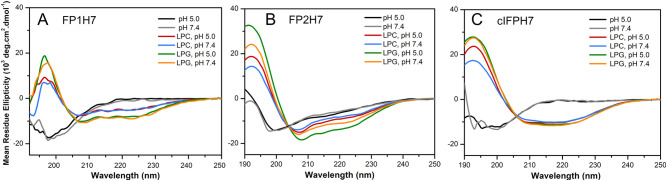
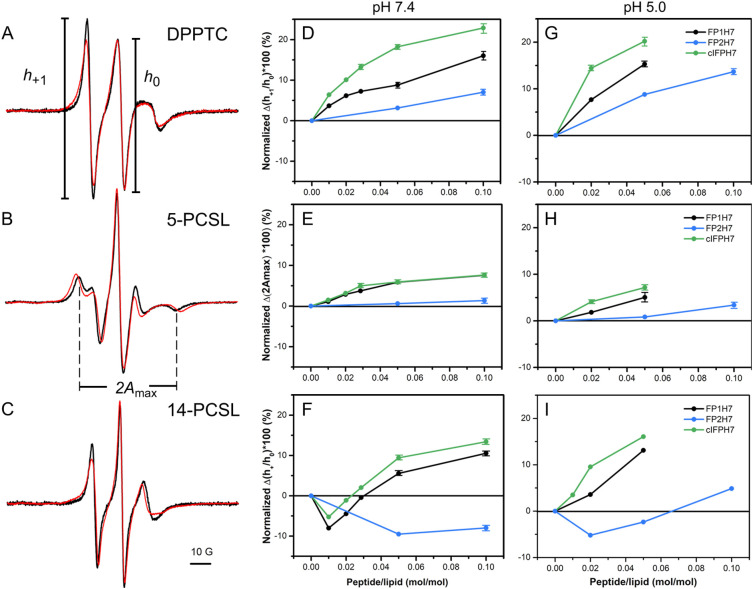
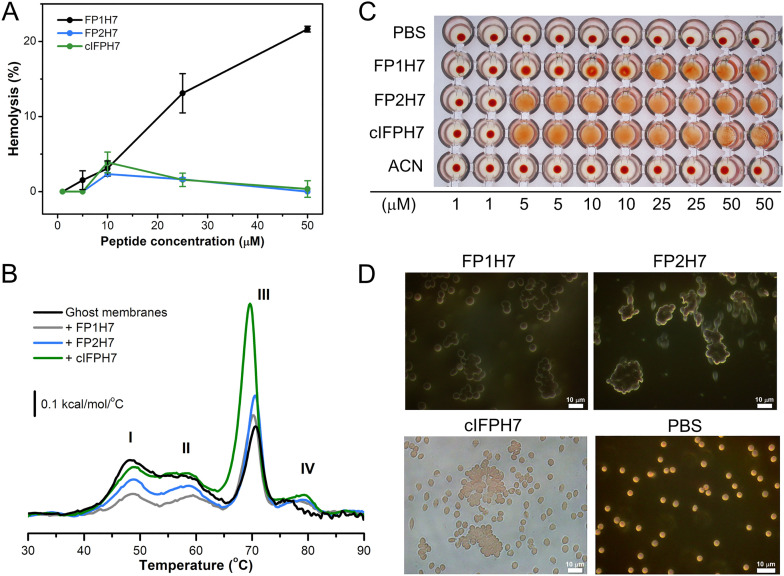
Fusion peptides (FP) are prominent hydrophobic segments of viral fusion proteins that play critical roles in viral entry. FPs interact with and insert into the host lipid membranes, triggering conformational changes in the viral protein that leads to the viral-cell fusion. Multiple membrane-active domains from the severe acute respiratory syndrome (SARS) coronavirus (CoV) spike protein have been reported to act as the functional fusion peptide such as the peptide sequence located between the S1/S2 and S2' cleavage sites (FP1), the S2'-adjacent fusion peptide domain (FP2), and the internal FP sequence (cIFP). Using a combined biophysical approach, we demonstrated that the α-helical coiled-coil-forming internal cIFP displayed the highest membrane fusion and permeabilizing activities along with membrane ordering effect in phosphatidylcholine (PC)/phosphatidylglycerol (PG) unilamellar vesicles compared to the other two N-proximal fusion peptide counterparts. While the FP1 sequence displayed intermediate membranotropic activities, the well-conserved FP2 peptide was substantially less effective in promoting fusion, leakage, and membrane ordering in PC/PG model membranes. Furthermore, Ca2+ did not enhance the FP2-induced lipid mixing activity in PC/phosphatidylserine/cholesterol lipid membranes, despite its strong erythrocyte membrane perturbation. Nonetheless, we found that the three putative SARS-CoV membrane-active fusion peptide sequences here studied altered the physical properties of model and erythrocyte membranes to different extents. The importance of the distinct membranotropic and biological activities of all SARS-CoV fusion peptide domains and the pronounced effect of the internal fusion peptide sequence to the whole spike-mediated membrane fusion process are discussed.
Keywords: COVID; Fusion peptide; Lipid-protein interaction; Membrane protein; SARS-CoV; SARS-CoV-2; Viral fusion.
Copyright © 2021 Elsevier B.V. All rights reserved.
Conflict of interest statement
ANTONIO J C FILHO reports financial support and equipment, drugs, or supplies were provided by State of Sao Paulo Research Foundation.
Figures
References
-
- Rota P.A., Oberste M.S., Monroe S.S., Nix W.A., Campagnoli R., Icenogle J.P., Bankamp B., Maher K., Chen M., Tong S., Tamin A., Lowe L., Frace M., Derisi J.L., Chen Q., Wang D., Erdman D.D., Peret T.C.T., Burns C., Ksiazek T.G., Rollin P.E., Sanchez A., Liffick S., Holloway B., Limor J., Mccaustland K., Olsen-rasmussen M., Fouchier R., Osterhaus A.D.M.E., Drosten C., Pallansch M.A., Anderson L.J., Bellini W.J. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science (80- ) 2003;300:1394–1399. - PubMed
-
- C.N.C.I. and R. Team. Zhu N., Zhang D., Wang W., Li X., Yang B., Song J., Zhao X., Huang B., Shi W., Lu R., Niu P., Zhan F., Ma X., Wang D., Xu W., Wu G., Gao G.F., Tan W. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020;382:727–733. doi: 10.1056/NEJMoa2001017. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
