

U5 snRNA Interactions With Exons Ensure Splicing Precision
- PMID: 34276781
- PMCID: PMC8283771
- DOI: 10.3389/fgene.2021.676971
U5 snRNA Interactions With Exons Ensure Splicing Precision
Abstract
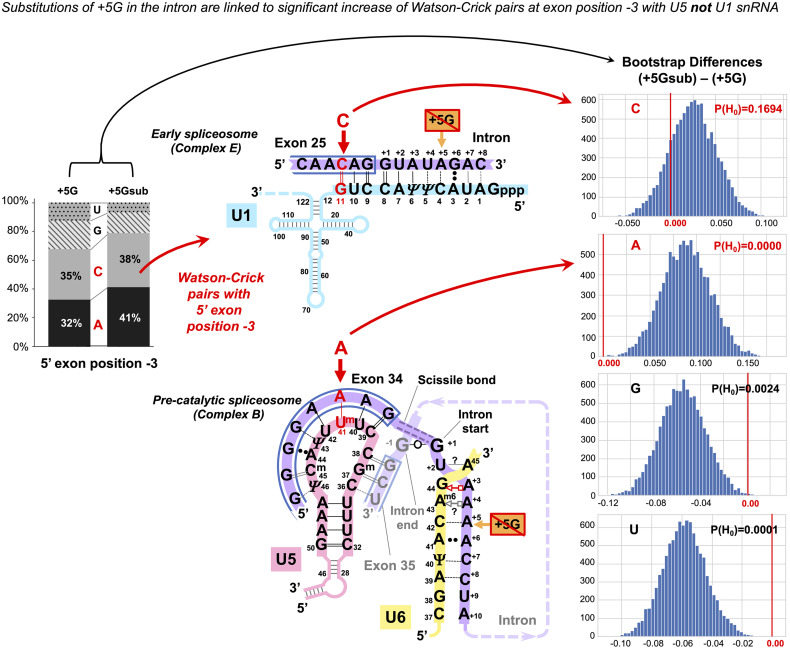
Imperfect conservation of human pre-mRNA splice sites is necessary to produce alternative isoforms. This flexibility is combined with the precision of the message reading frame. Apart from intron-termini GU_AG and the branchpoint A, the most conserved are the exon-end guanine and +5G of the intron start. Association between these guanines cannot be explained solely by base-pairing with U1 snRNA in the early spliceosome complex. U6 succeeds U1 and pairs +5G in the pre-catalytic spliceosome, while U5 binds the exon end. Current U5 snRNA reconstructions by CryoEM cannot explain the conservation of the exon-end G. Conversely, human mutation analyses show that guanines of both exon termini can suppress splicing mutations. Our U5 hypothesis explains the mechanism of splicing precision and the role of these conserved guanines in the pre-catalytic spliceosome. We propose: (1) optimal binding register for human exons and U5-the exon junction positioned at U5Loop1 C39|C38; (2) common mechanism for base-pairing of human U5 snRNA with diverse exons and bacterial Ll.LtrB intron with new loci in retrotransposition-guided by base pair geometry; and (3) U5 plays a significant role in specific exon recognition in the pre-catalytic spliceosome. Statistical analyses showed increased U5 Watson-Crick pairs with the 5'exon in the absence of +5G at the intron start. In 5'exon positions -3 and -5, this effect is specific to U5 snRNA rather than U1 snRNA of the early spliceosome. Increased U5 Watson-Crick pairs with 3'exon position +1 coincide with substitutions of the conserved -3C at the intron 3'end. Based on mutation and X-ray evidence, we propose that -3C pairs with U2 G31 juxtaposing the branchpoint and the 3'intron end. The intron-termini pair, formed in the pre-catalytic spliceosome to be ready for transition after branching, and the early involvement of the 3'intron end ensure that the 3'exon contacts U5 in the pre-catalytic complex. We suggest that splicing precision is safeguarded cooperatively by U5, U6, and U2 snRNAs that stabilize the pre-catalytic complex by Watson-Crick base pairing. In addition, our new U5 model explains the splicing effect of exon-start +1G mutations: U5 Watson-Crick pairs with exon +2C/+3G strongly promote exon inclusion. We discuss potential applications for snRNA therapeutics and gene repair by reverse splicing.
Keywords: RNA base pair geometry; U1 snRNA; U2 snRNA; U5 snRNA; U6 snRNA; group II intron retrotransposition; splice sites; splicing mutations.
Copyright © 2021 Artemyeva-Isman and Porter.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures













References
-
- Bai R., Yan C., Wan R., Lei J., Shi Y. (2017). Structure of the post-catalytic spliceosome from Saccharomyces cerevisiae. Cell 171 1589–1598.e8. - PubMed
LinkOut - more resources
Full Text Sources