

A Gut-Restricted Lithocholic Acid Analog as an Inhibitor of Gut Bacterial Bile Salt Hydrolases
- PMID: 34279901
- PMCID: PMC9013266
- DOI: 10.1021/acschembio.1c00192
A Gut-Restricted Lithocholic Acid Analog as an Inhibitor of Gut Bacterial Bile Salt Hydrolases
Abstract
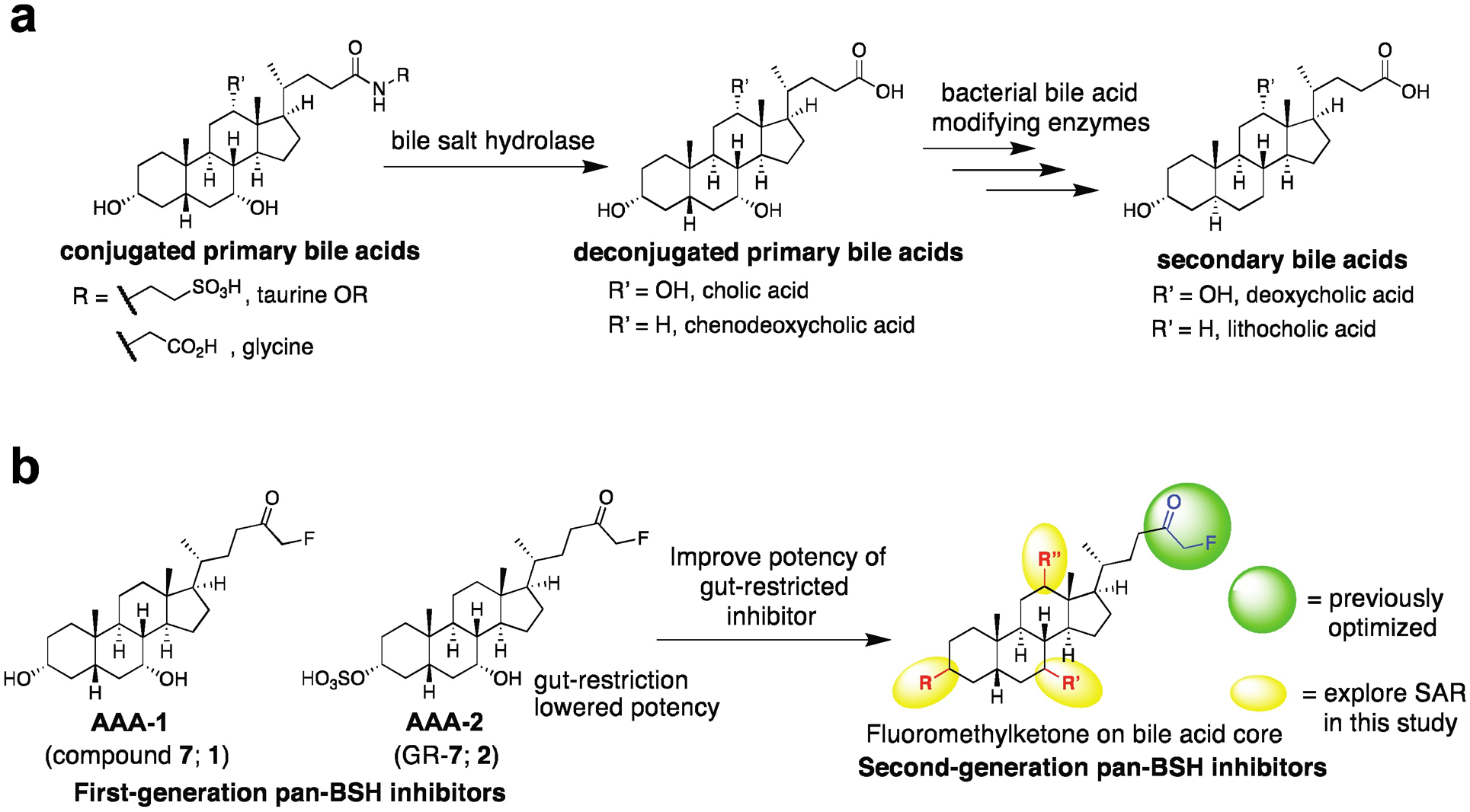
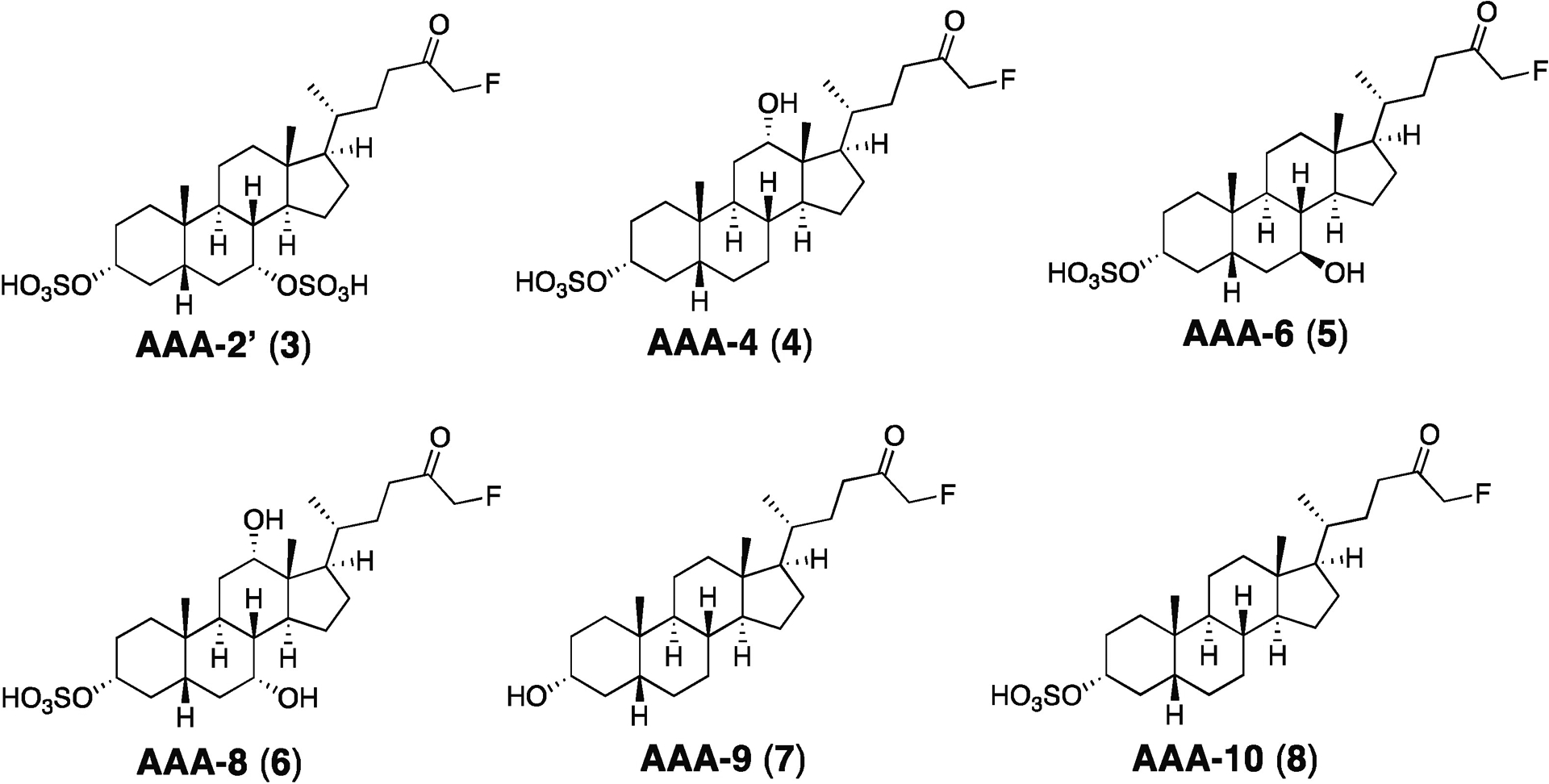
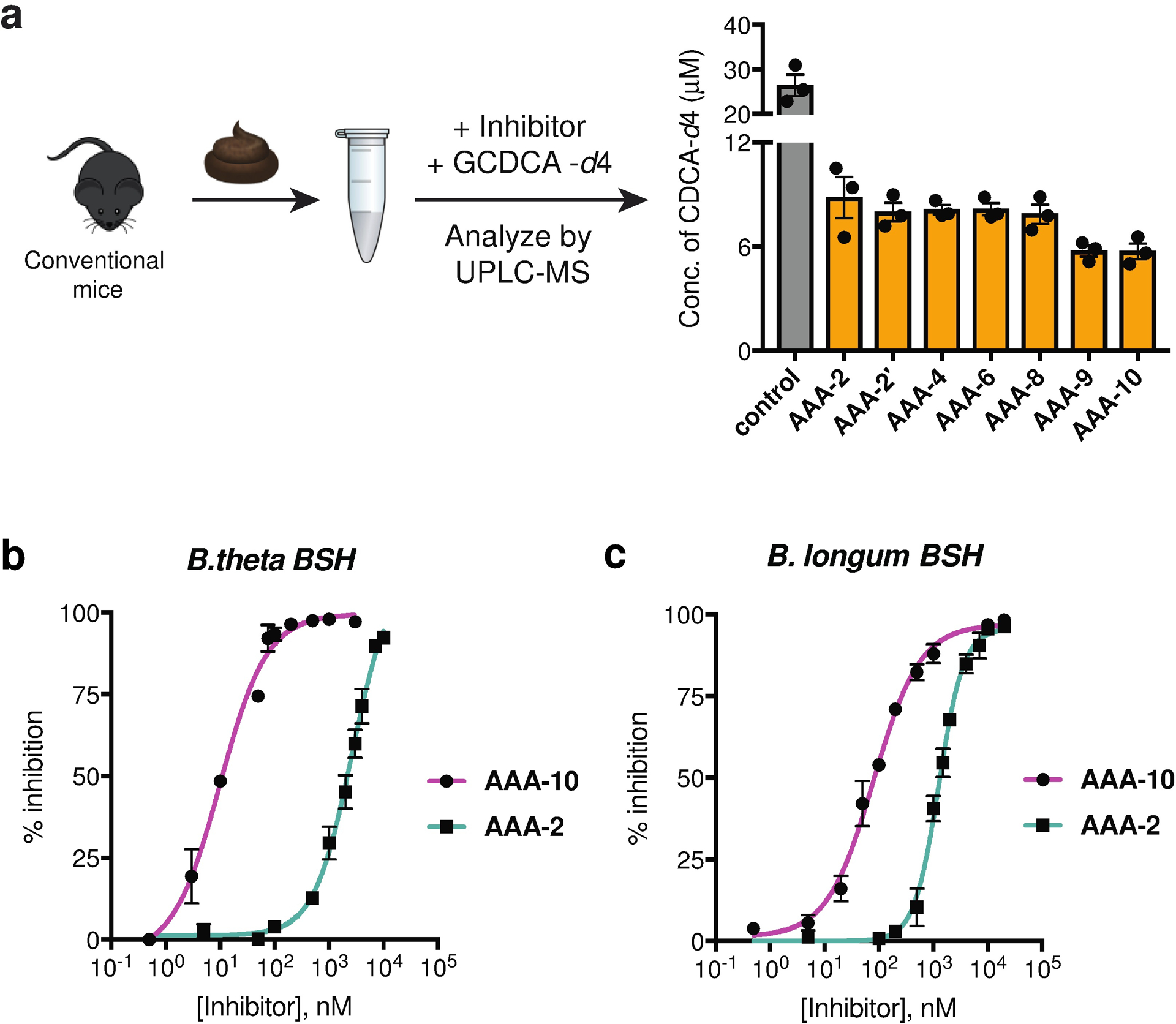
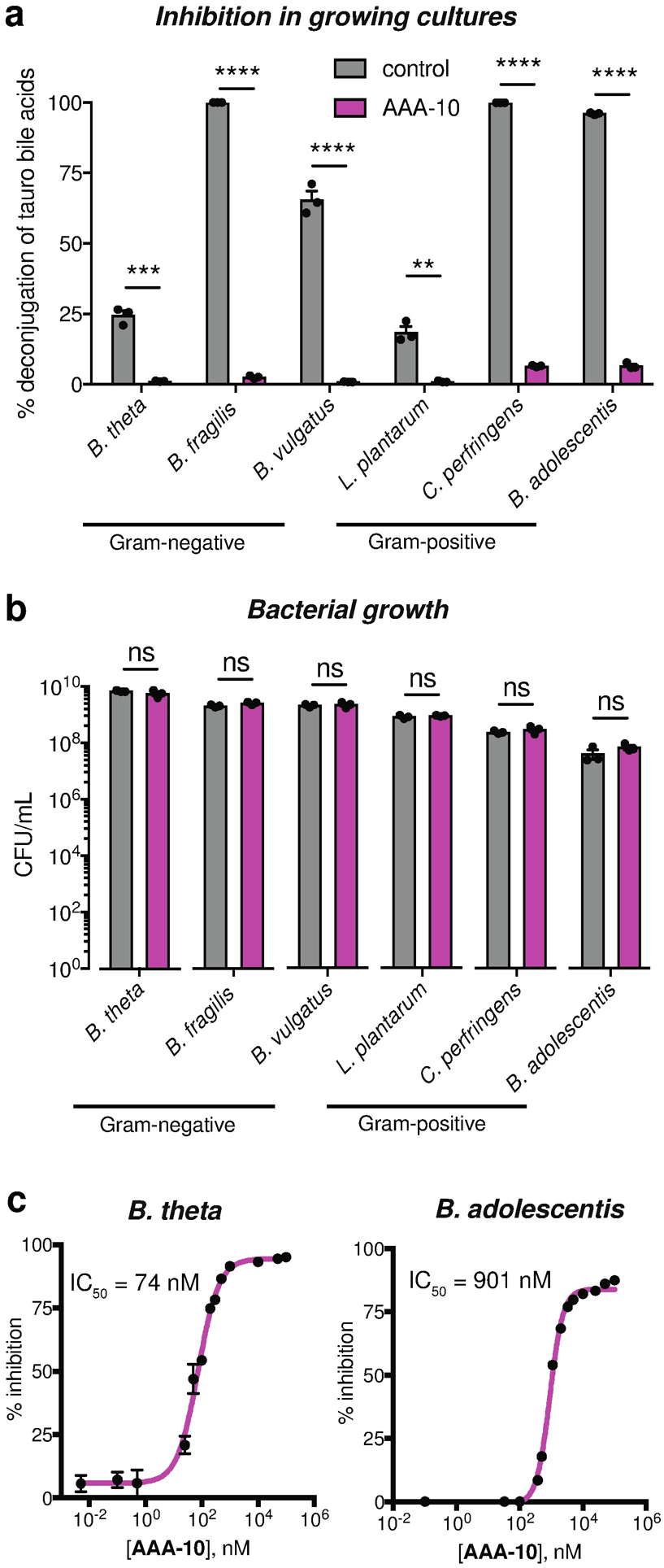
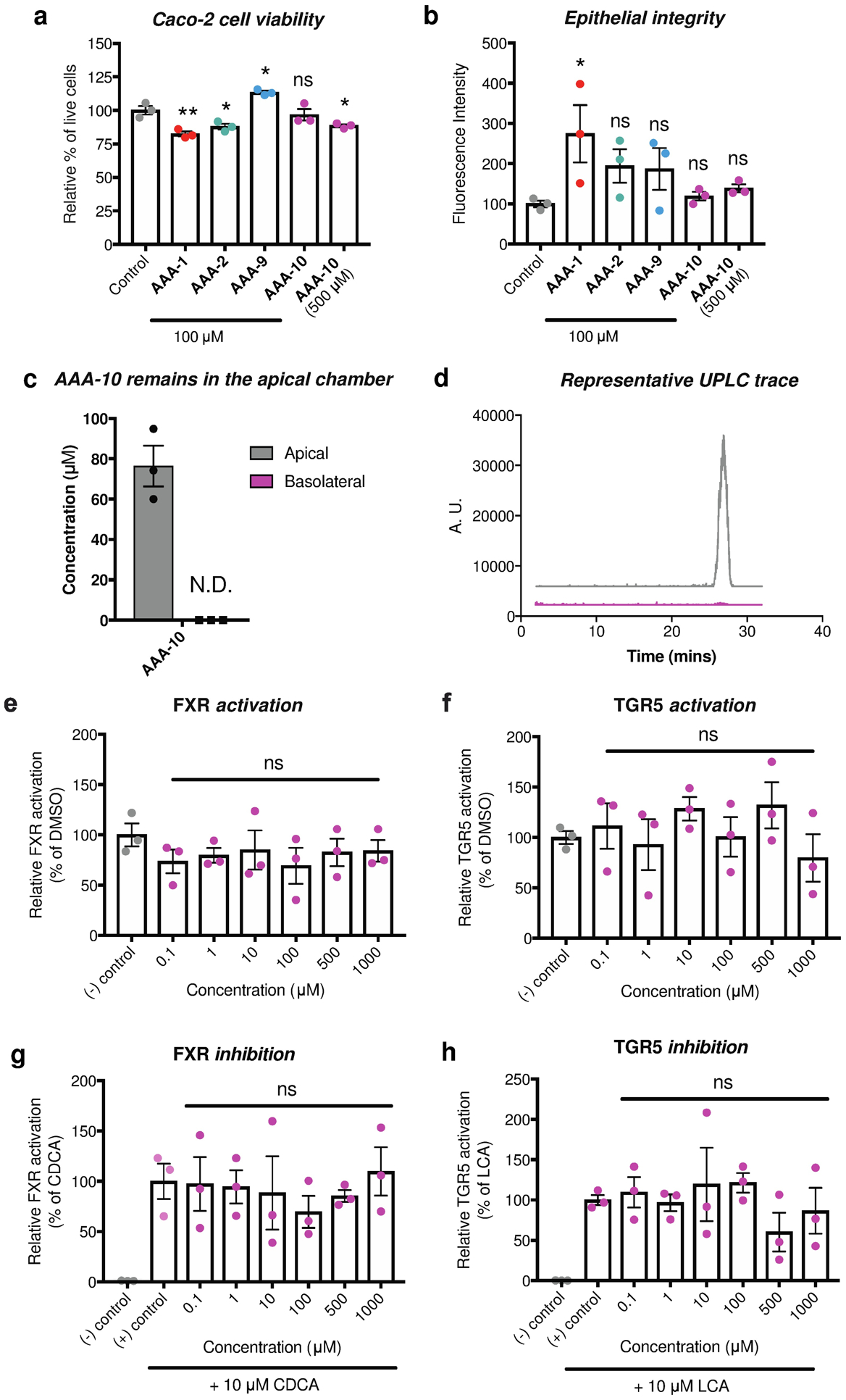
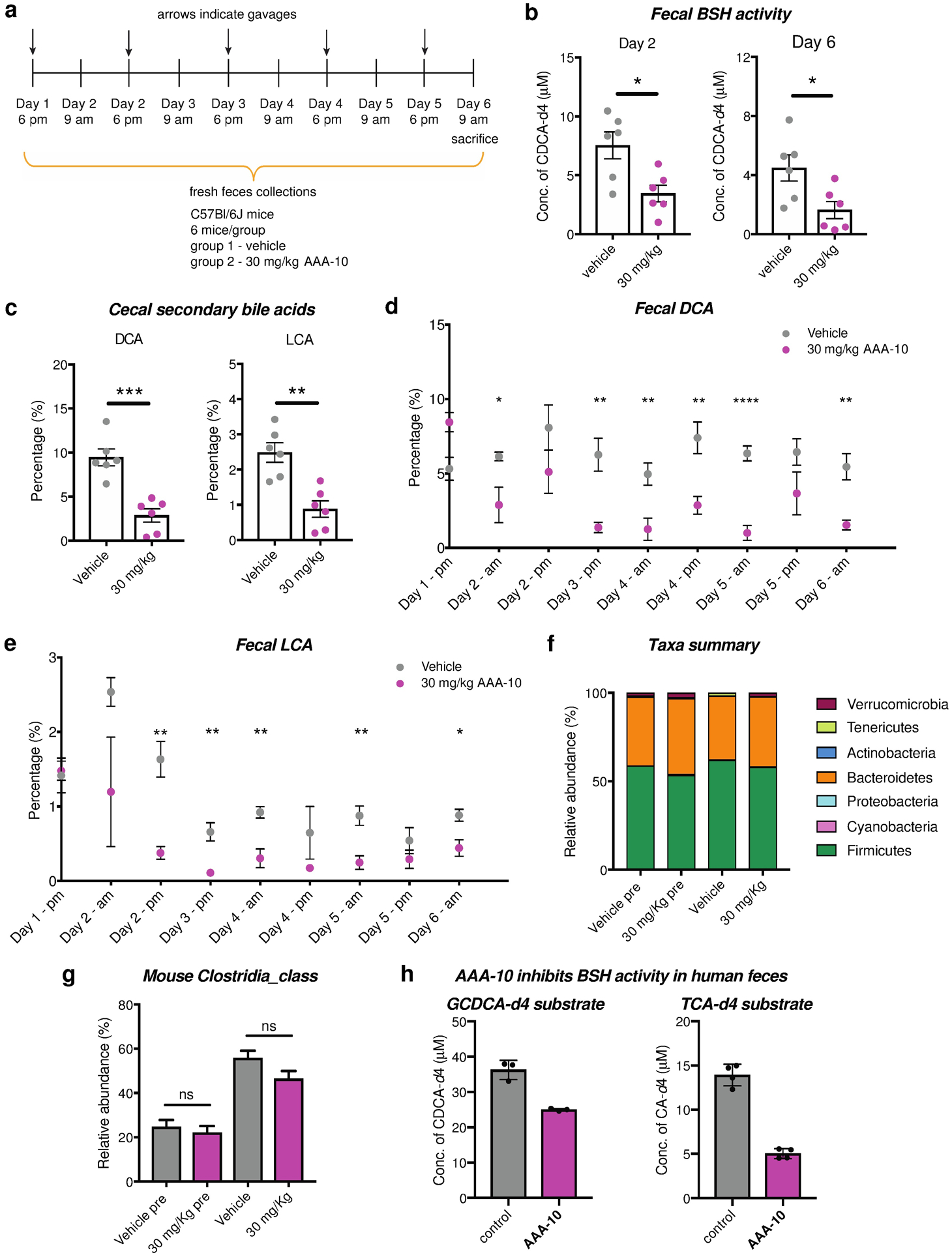
Bile acids play crucial roles in host physiology by acting both as detergents that aid in digestion and as signaling molecules that bind to host receptors. Gut bacterial bile salt hydrolase (BSH) enzymes perform the gateway reaction leading to the conversion of host-produced primary bile acids into bacterially modified secondary bile acids. Small molecule probes that target BSHs will help elucidate the causal roles of these metabolites in host physiology. We previously reported the development of a covalent BSH inhibitor with low gut permeability. Here, we build on our previous findings and describe the development of a second-generation gut-restricted BSH inhibitor with enhanced potency, reduced off-target effects, and durable in vivo efficacy. Structure-activity relationship (SAR) studies focused on the bile acid core identified a compound, AAA-10, containing a C3-sulfonated lithocholic acid scaffold and an alpha-fluoromethyl ketone warhead as a potent pan-BSH inhibitor. This compound inhibits BSH activity in mouse and human fecal slurry, bacterial cultures, and purified BSH proteins and displays reduced toxicity against mammalian cells compared to first generation compounds. Oral administration of AAA-10 to wild-type mice for 5 days resulted in a decrease in the abundance of the secondary bile acids deoxycholic acid (DCA) and lithocholic acid (LCA) in the mouse GI tract with low systemic exposure of AAA-10, demonstrating that AAA-10 is an effective tool for inhibiting BSH activity and modulating bile acid pool composition in vivo.
Conflict of interest statement
The authors declare the following competing financial interest(s): A. Sloan Devlin is an ad hoc consultant for Takeda Pharmaceuticals and Axial Therapeutics. The other authors declare that no competing interests exist.
Figures
References
-
- Roberts AB, Gu X, Buffa JA, Hurd AG, Wang Z, Zhu W, Gupta N, Skye SM, Cody DB, Levison BS, Barrington WT, Russell MW, Reed JM, Duzan A, Lang JM, Fu X, Li L, Myers AJ, Rachakonda S, DiDonato JA, Brown JM, Gogonea V, Lusis AJ, Garcia-Garcia JC, and Hazen SL (2018) Development of a gut microbe-targeted nonlethal therapeutic to inhibit thrombosis potential. Nat. Med 24, 1407–1417. - PMC - PubMed
-
- Ridlon JM, Kang D-J, and Hylemon PB (2006) Bile salt biotransformations by human intestinal bacteria. J. Lipid Res 47, 241–259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
