

Multi-scale simulations of the T cell receptor reveal its lipid interactions, dynamics and the arrangement of its cytoplasmic region
- PMID: 34280187
- PMCID: PMC8321403
- DOI: 10.1371/journal.pcbi.1009232
Multi-scale simulations of the T cell receptor reveal its lipid interactions, dynamics and the arrangement of its cytoplasmic region
Abstract
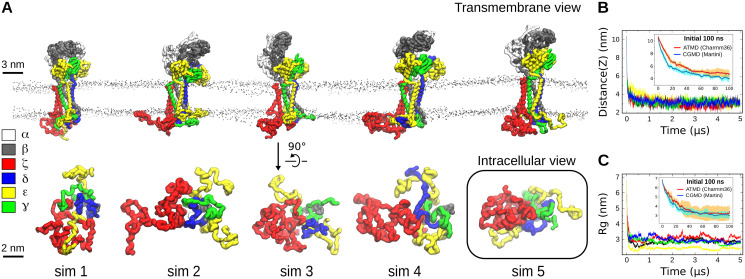
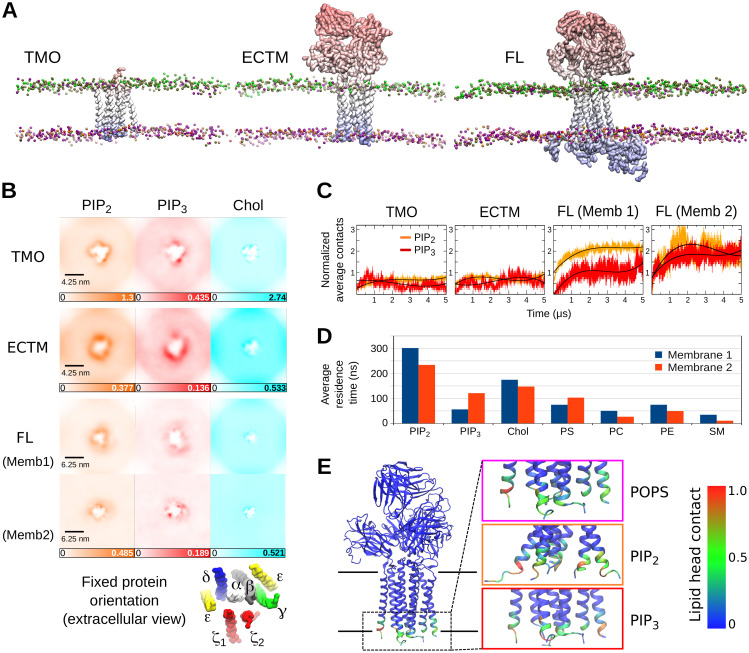
The T cell receptor (TCR-CD3) initiates T cell activation by binding to peptides of Major Histocompatibility Complexes (pMHC). The TCR-CD3 topology is well understood but the arrangement and dynamics of its cytoplasmic tails remains unknown, limiting our grasp of the signalling mechanism. Here, we use molecular dynamics simulations and modelling to investigate the entire TCR-CD3 embedded in a model membrane. Our study demonstrates conformational changes in the extracellular and transmembrane domains, and the arrangement of the TCR-CD3 cytoplasmic tails. The cytoplasmic tails formed highly interlaced structures while some tyrosines within the immunoreceptor tyrosine-based activation motifs (ITAMs) penetrated the hydrophobic core of the membrane. Interactions between the cytoplasmic tails and phosphatidylinositol phosphate lipids in the inner membrane leaflet led to the formation of a distinct anionic lipid fingerprint around the TCR-CD3. These results increase our understanding of the TCR-CD3 dynamics and the importance of membrane lipids in regulating T cell activation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Peptide-MHC (pMHC) binding to a human antiviral T cell receptor induces long-range allosteric communication between pMHC- and CD3-binding sites.J Biol Chem. 2018 Oct 12;293(41):15991-16005. doi: 10.1074/jbc.RA118.003832. Epub 2018 Aug 22. J Biol Chem. 2018. PMID: 30135211 Free PMC article.
-
Ca2+ regulates T-cell receptor activation by modulating the charge property of lipids.Nature. 2013 Jan 3;493(7430):111-5. doi: 10.1038/nature11699. Epub 2012 Dec 2. Nature. 2013. PMID: 23201688
-
Structural variability and concerted motions of the T cell receptor - CD3 complex.Elife. 2021 Sep 7;10:e67195. doi: 10.7554/eLife.67195. Elife. 2021. PMID: 34490842 Free PMC article.
-
The TCR/CD3 complex: molecular interactions in a changing structure.Arch Immunol Ther Exp (Warsz). 2002;50(4):263-72. Arch Immunol Ther Exp (Warsz). 2002. PMID: 12371622 Review.
-
Organization of proximal signal initiation at the TCR:CD3 complex.Immunol Rev. 2009 Nov;232(1):7-21. doi: 10.1111/j.1600-065X.2009.00843.x. Immunol Rev. 2009. PMID: 19909352 Free PMC article. Review.
Cited by
-
A dynamic biomimetic model of the membrane-bound CD4-CD3-TCR complex during pMHC disengagement.Biophys J. 2023 Aug 8;122(15):3133-3145. doi: 10.1016/j.bpj.2023.06.018. Epub 2023 Jun 28. Biophys J. 2023. PMID: 37381600 Free PMC article.
-
Molecular dynamics simulations reveal membrane lipid interactions of the full-length lymphocyte specific kinase (Lck).Sci Rep. 2022 Dec 7;12(1):21121. doi: 10.1038/s41598-022-25603-6. Sci Rep. 2022. PMID: 36476673 Free PMC article.
-
In Silico Assessment of the Lipid Fingerprint Signature of ATP2, the Essential P4-ATPase of Malaria Parasites.Membranes (Basel). 2022 Jul 12;12(7):702. doi: 10.3390/membranes12070702. Membranes (Basel). 2022. PMID: 35877905 Free PMC article.
-
Mechanical force matters in early T cell activation.Proc Natl Acad Sci U S A. 2024 Sep 10;121(37):e2404748121. doi: 10.1073/pnas.2404748121. Epub 2024 Sep 6. Proc Natl Acad Sci U S A. 2024. PMID: 39240966 Free PMC article.
-
Recent advances in understanding TCR signaling: a synaptic perspective.Fac Rev. 2023 Oct 16;12:25. doi: 10.12703/r/12-25. eCollection 2023. Fac Rev. 2023. PMID: 37900153 Free PMC article. Review.
