

Structural basis for template switching by a group II intron-encoded non-LTR-retroelement reverse transcriptase
- PMID: 34280434
- PMCID: PMC8363836
- DOI: 10.1016/j.jbc.2021.100971
Structural basis for template switching by a group II intron-encoded non-LTR-retroelement reverse transcriptase
Abstract
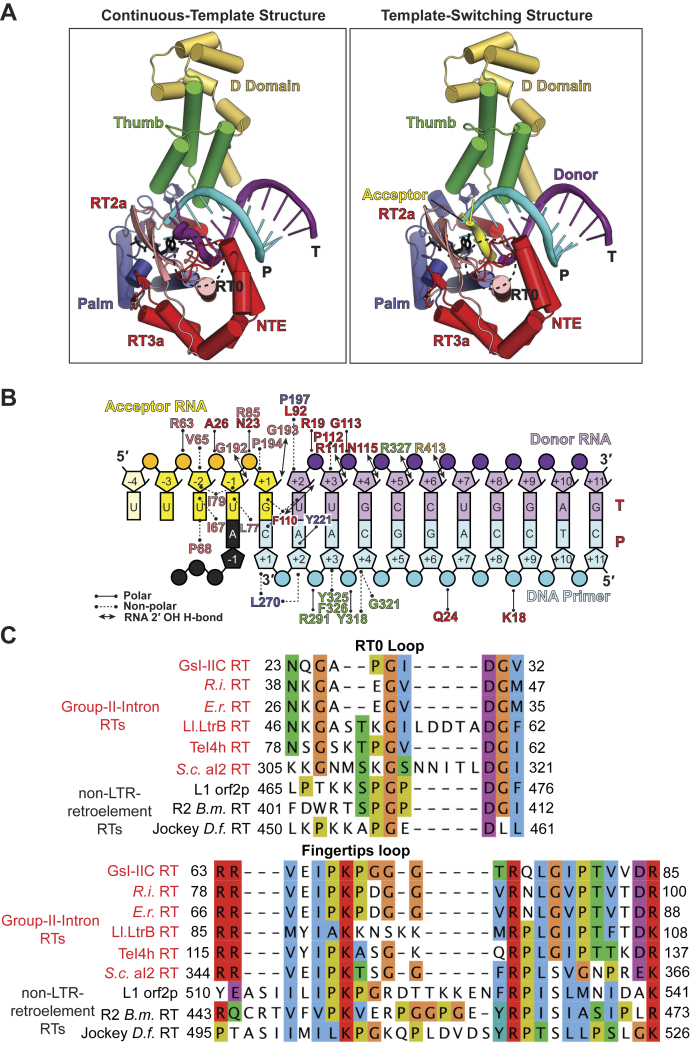
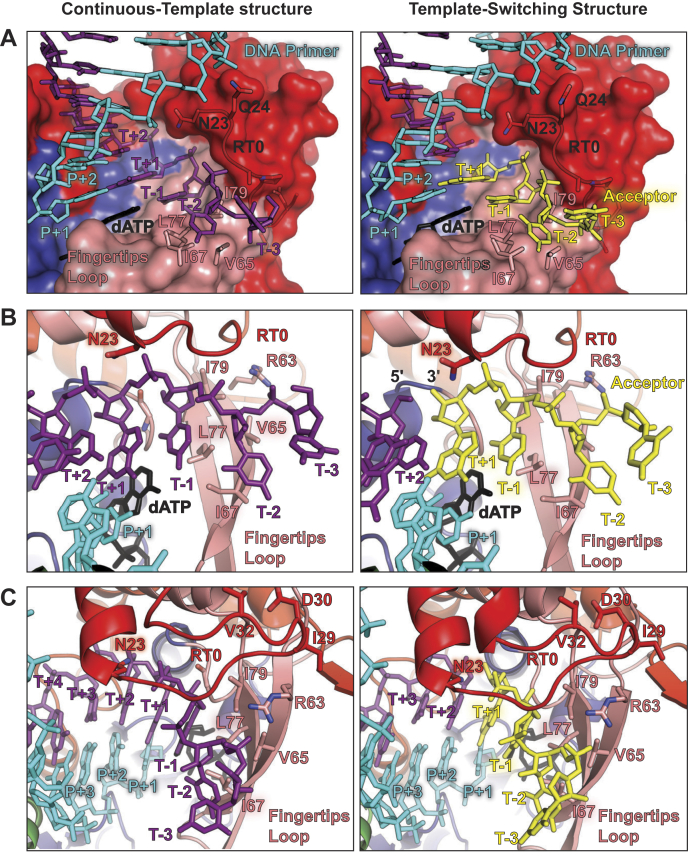
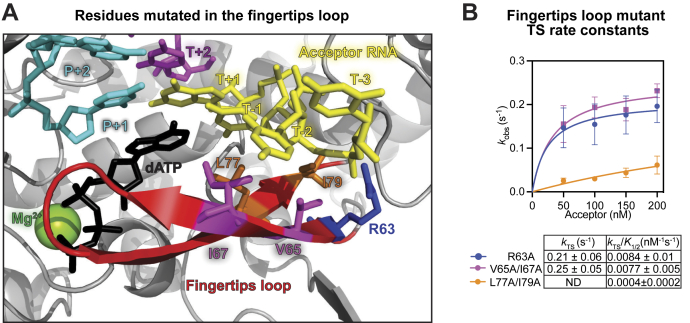
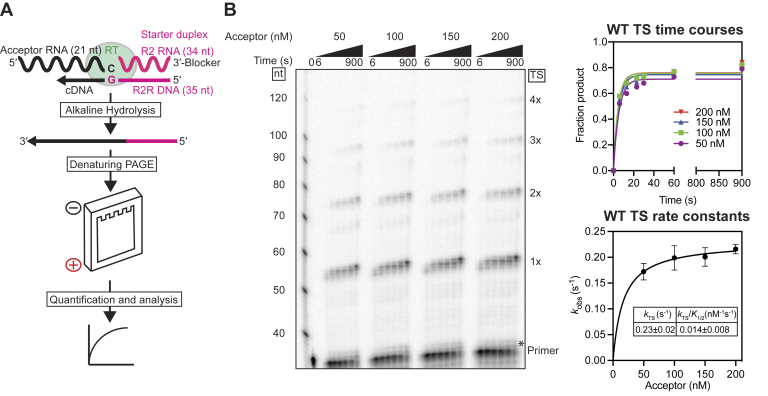
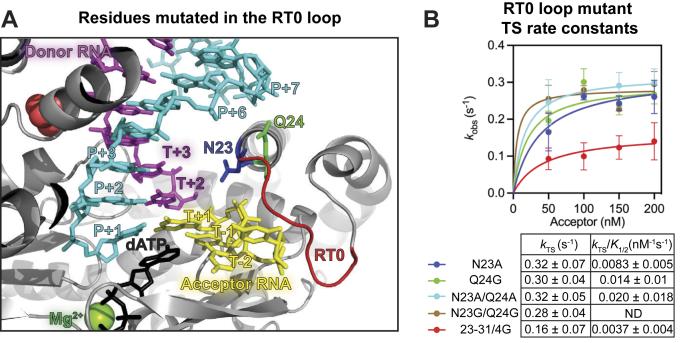
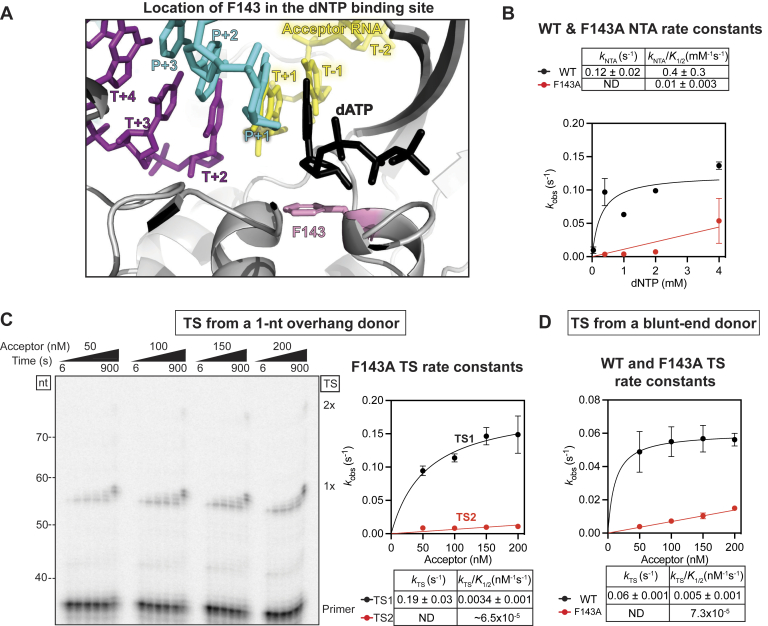
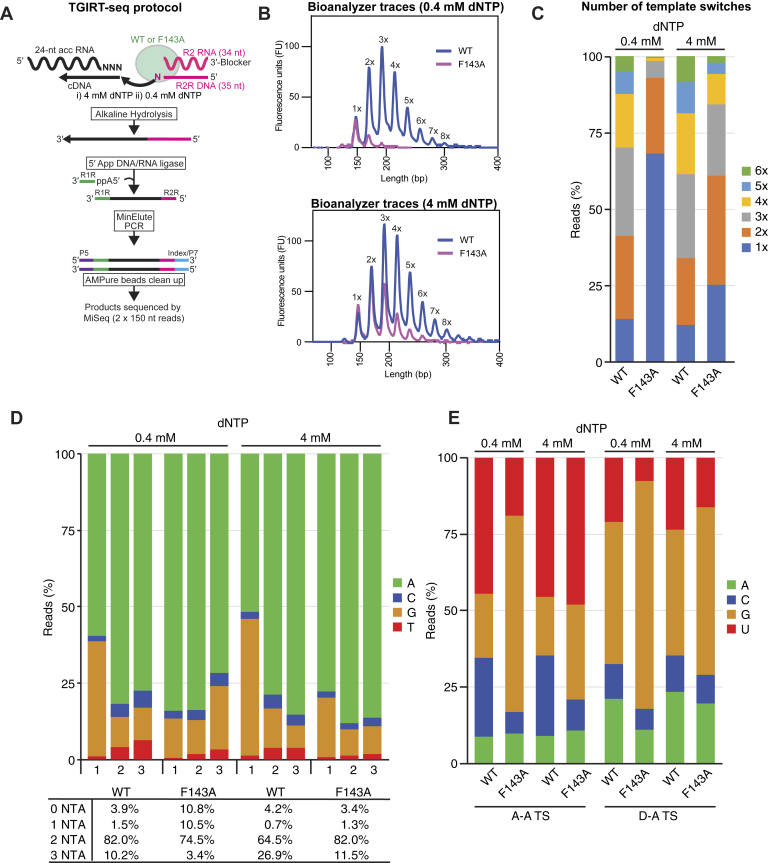
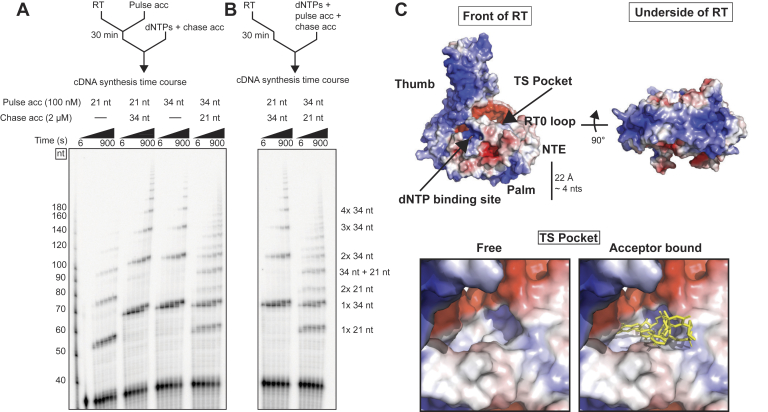
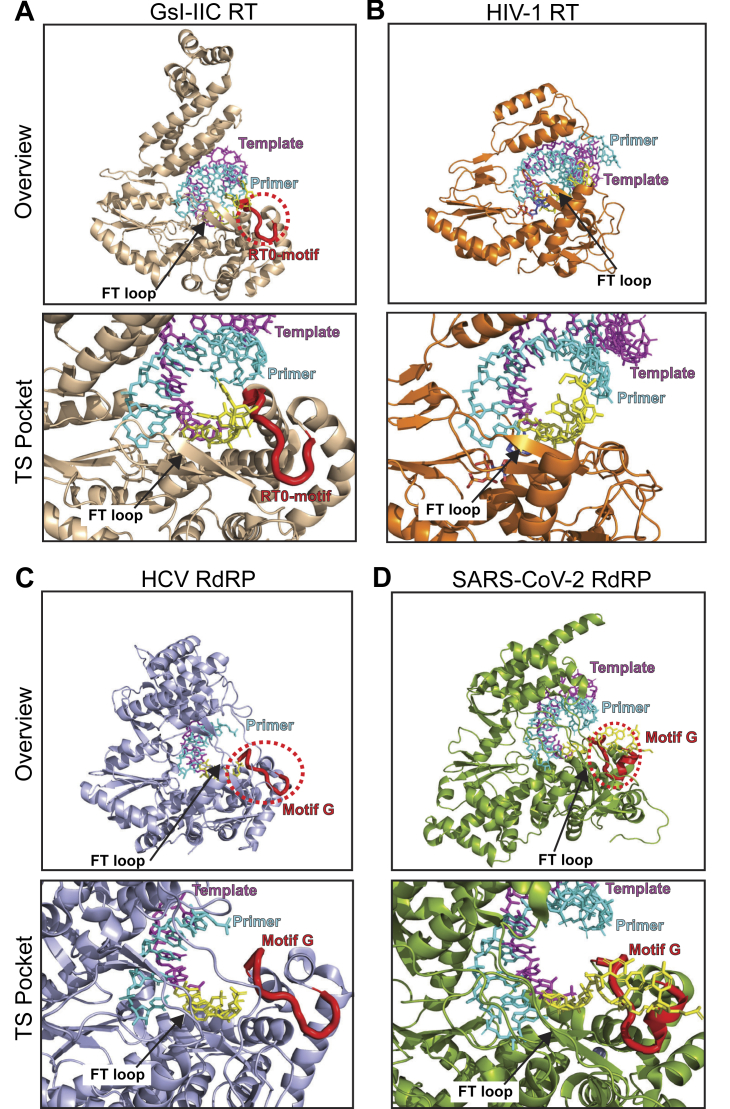
Reverse transcriptases (RTs) can switch template strands during complementary DNA synthesis, enabling them to join discontinuous nucleic acid sequences. Template switching (TS) plays crucial roles in retroviral replication and recombination, is used for adapter addition in RNA-Seq, and may contribute to retroelement fitness by increasing evolutionary diversity and enabling continuous complementary DNA synthesis on damaged templates. Here, we determined an X-ray crystal structure of a TS complex of a group II intron RT bound simultaneously to an acceptor RNA and donor RNA template-DNA primer heteroduplex with a 1-nt 3'-DNA overhang. The structure showed that the 3' end of the acceptor RNA binds in a pocket formed by an N-terminal extension present in non-long terminal repeat-retroelement RTs and the RT fingertips loop, with the 3' nucleotide of the acceptor base paired to the 1-nt 3'-DNA overhang and its penultimate nucleotide base paired to the incoming dNTP at the RT active site. Analysis of structure-guided mutations identified amino acids that contribute to acceptor RNA binding and a phenylalanine residue near the RT active site that mediates nontemplated nucleotide addition. Mutation of the latter residue decreased multiple sequential template switches in RNA-Seq. Our results provide new insights into the mechanisms of TS and nontemplated nucleotide addition by RTs, suggest how these reactions could be improved for RNA-Seq, and reveal common structural features for TS by non-long terminal repeat-retroelement RTs and viral RNA-dependent RNA polymerases.
Keywords: RNA virus; RNA-dependent RNA polymerase; X-ray crystallography; high-throughput RNA-Seq; retrovirus; thermostable group II intron reverse transcriptase; transcriptomics.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest Thermostable group II intron RT enzymes and methods for their use are the subject of patents and patent applications that have been licensed by the University of Texas and East Tennessee State University to InGex, LLC. A. M. Lambowitz, some former and present members of the Lambowitz laboratory, and the University of Texas are minority equity holders in InGex, LLC, and receive royalty payments from the sale of TGIRT enzymes and kits employing TGIRT TS activity and from the sublicensing of intellectual property to other companies. All other authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Coffin J.M. Structure, replication, and recombination of retrovirus genomes: Some unifying hypotheses. J. Gen. Virol. 1979;42:1–26. - PubMed
-
- Shih C., Yang C.C., Choijilsuren G., Chang C.H., Liou A.T. Hepatitis B virus. Trends Microbiol. 2018;26:386–387. - PubMed
-
- Martin-Alonso S., Frutos-Beltran E., Menendez-Arias L. Reverse transcriptase: From transcriptomics to genome editing. Trends Biotechnol. 2021;39:194–210. - PubMed
-
- Inouye S., Hsu M.Y., Eagle S., Inouye M. Reverse transcriptase associated with the biosynthesis of the branched RNA-linked msDNA in Myxococcus xanthus. Cell. 1989;56:709–717. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
