

Chronic Orofacial Pain: Models, Mechanisms, and Genetic and Related Environmental Influences
- PMID: 34281164
- PMCID: PMC8268972
- DOI: 10.3390/ijms22137112
Chronic Orofacial Pain: Models, Mechanisms, and Genetic and Related Environmental Influences
Abstract
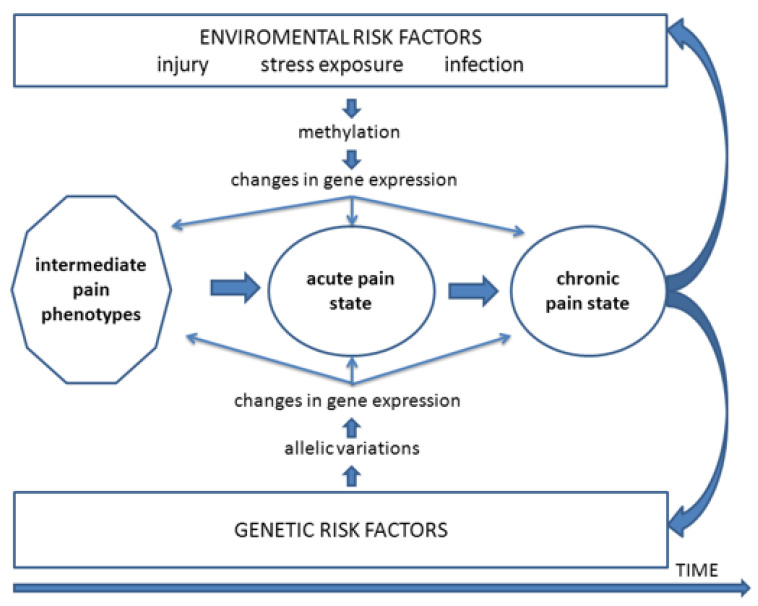
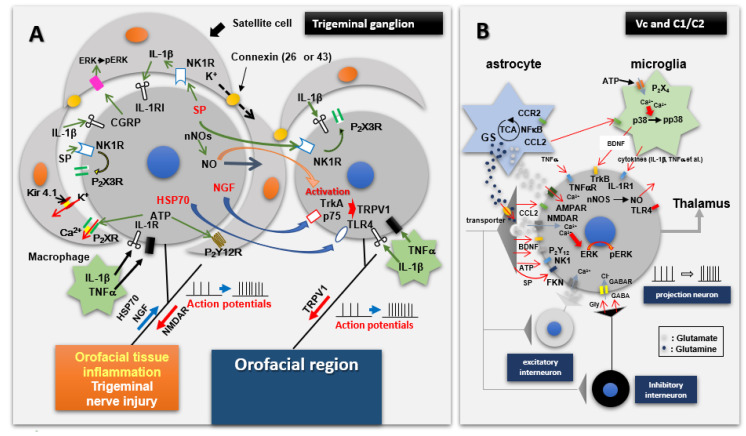
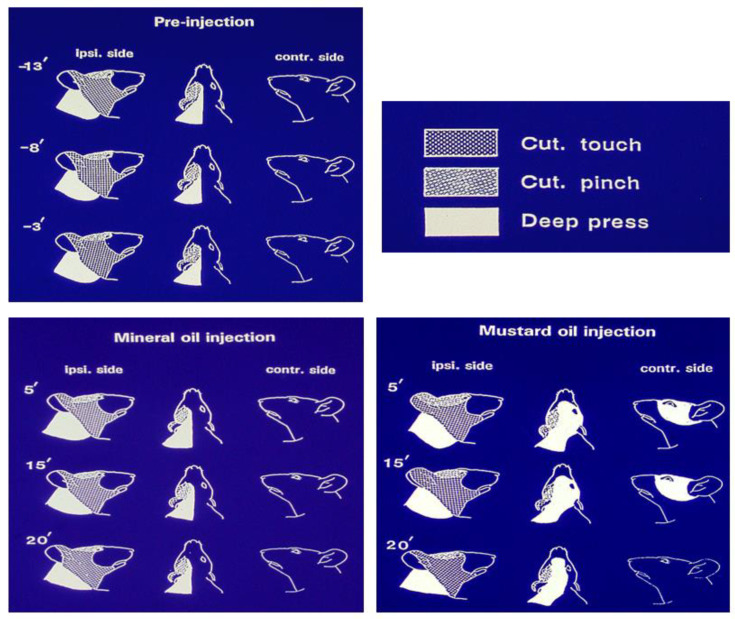
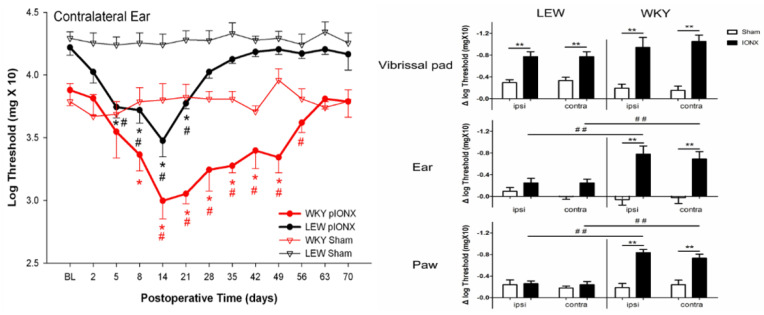
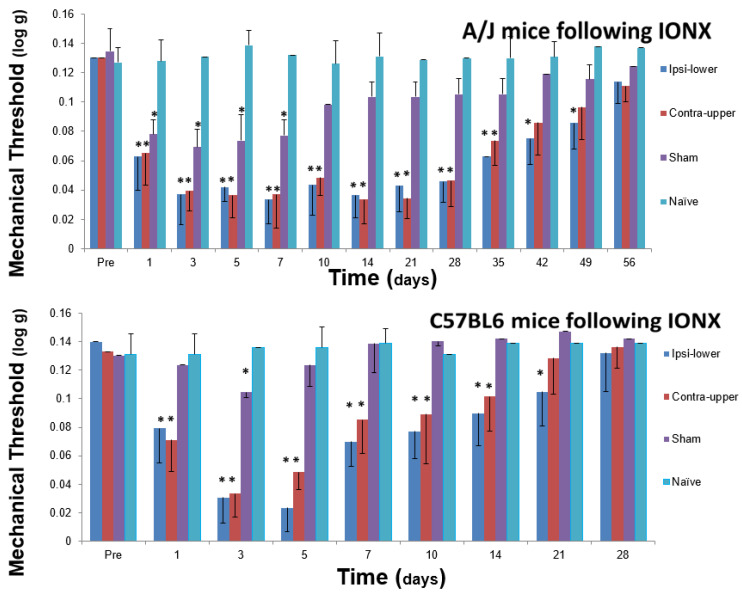
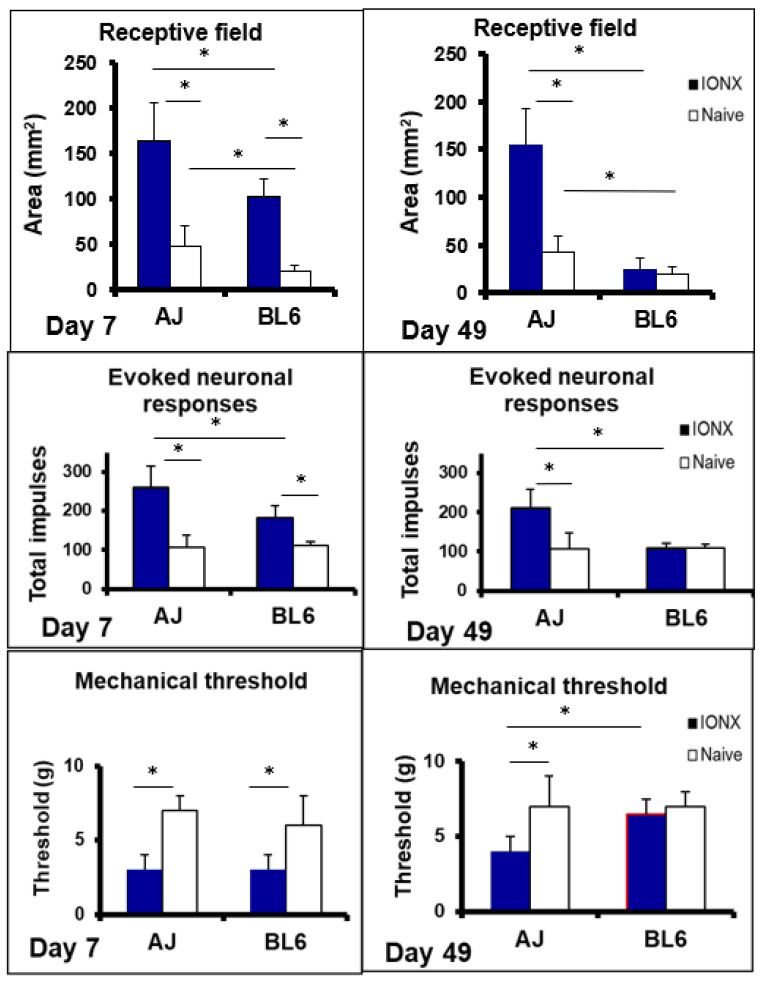
Chronic orofacial pain conditions can be particularly difficult to diagnose and treat because of their complexity and limited understanding of the mechanisms underlying their aetiology and pathogenesis. Furthermore, there is considerable variability between individuals in their susceptibility to risk factors predisposing them to the development and maintenance of chronic pain as well as in their expression of chronic pain features such as allodynia, hyperalgesia and extraterritorial sensory spread. The variability suggests that genetic as well as environmental factors may contribute to the development and maintenance of chronic orofacial pain. This article reviews these features of chronic orofacial pain, and outlines findings from studies in animal models of the behavioural characteristics and underlying mechanisms related to the development and maintenance of chronic orofacial pain and trigeminal neuropathic pain in particular. The review also considers the role of environmental and especially genetic factors in these models, focussing on findings of differences between animal strains in the features and underlying mechanisms of chronic pain. These findings are not only relevant to understanding underlying mechanisms and the variability between patients in the development, expression and maintenance of chronic orofacial pain, but also underscore the importance for considering the strain of the animal to model and explore chronic orofacial pain processes.
Keywords: animal models; environmental factors; genetic factors; orofacial pain; strain differences; trigeminal.
Conflict of interest statement
The author declares no conflict of interest.
Figures
References
-
- Arendt-Nielsen L., Graven-Nielsen T., Sessle B.J. Mechanisms underlying extraterritorial and widespread sensitization: From animal to chronic pain. In: Graven-Nielsen T., Arendt-Nielsen L., editors. Musculoskeletal Pain: Basic Mechanisms & Implications. Wolters Kluwer Health; Washington, DC, USA: 2015. pp. 417–436.
-
- Macfarlane T.V. Epidemiology of orofacial pain. In: Sessle B.J., editor. Orofacial Pain: Recent Advances in Assessment, Management, and Understanding of Mechanisms. IASP Press; Washington, DC, USA: 2014. pp. 33–52.
-
- Svensson P., Baad-Hansen L., Drangsholt M., Jaaskelainen S. Neurosensory testing for assessment, diagnosis, and prediction of orofacial pain. In: Sessle B.J., editor. Orofacial Pain: Recent Advances in Assessment, Management, and Understanding of Mechanisms. IASP Press; Washington, DC, USA: 2014. pp. 143–164.
-
- Slade G.D., Greenspan J.D., Fillingim R.B., Maixner W., Sharma S., Ohrbach R. Overlap of five chronic pain conditions: Temporomandibular disorders, headache, back pain, irritable bowel syndrome, and fibromyalgia. J. Oral Facial Pain Headache. 2020;34:s15–s28. doi: 10.11607/ofph.2581. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
