

Arabidopsis group C Raf-like protein kinases negatively regulate abscisic acid signaling and are direct substrates of SnRK2
- PMID: 34282011
- PMCID: PMC8325330
- DOI: 10.1073/pnas.2100073118
Arabidopsis group C Raf-like protein kinases negatively regulate abscisic acid signaling and are direct substrates of SnRK2
Abstract
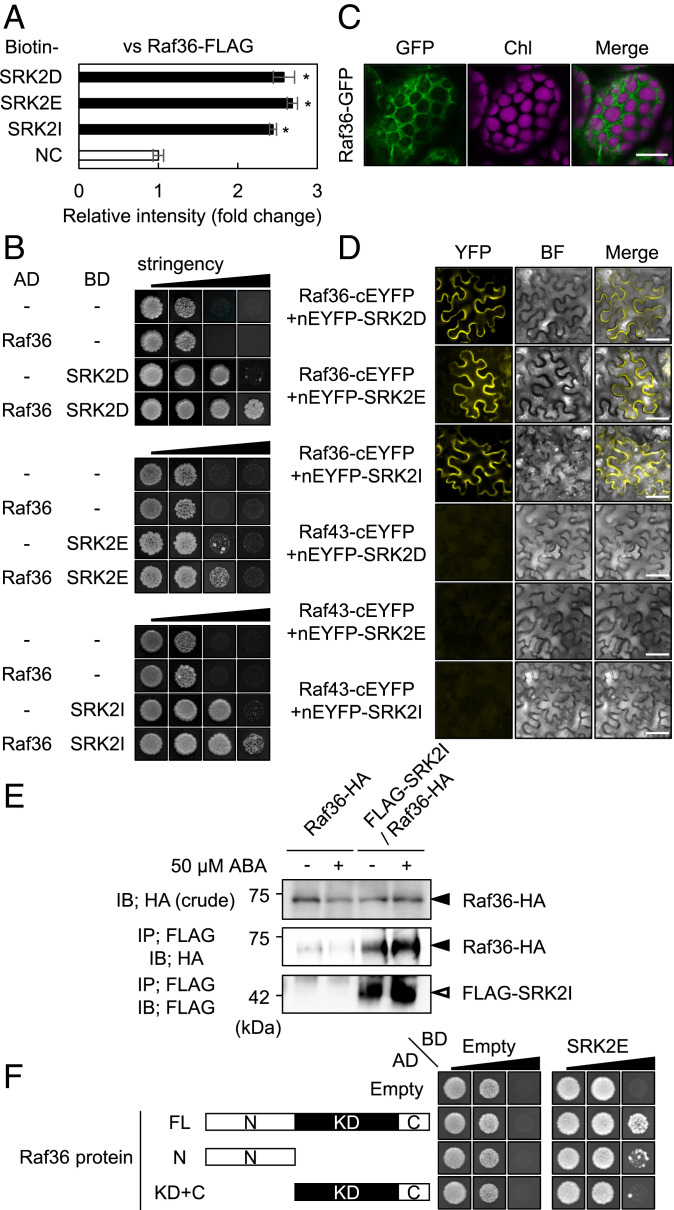
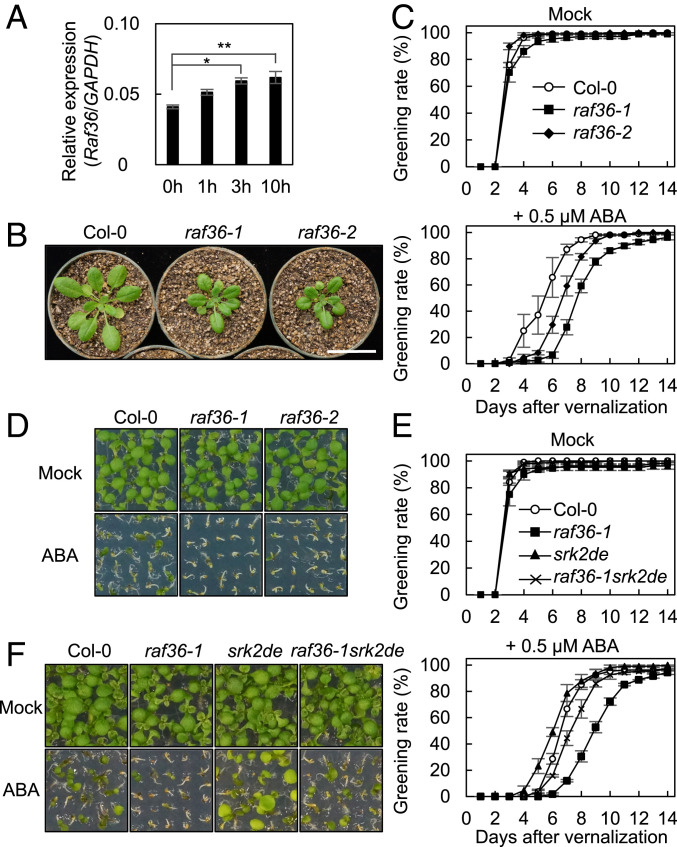
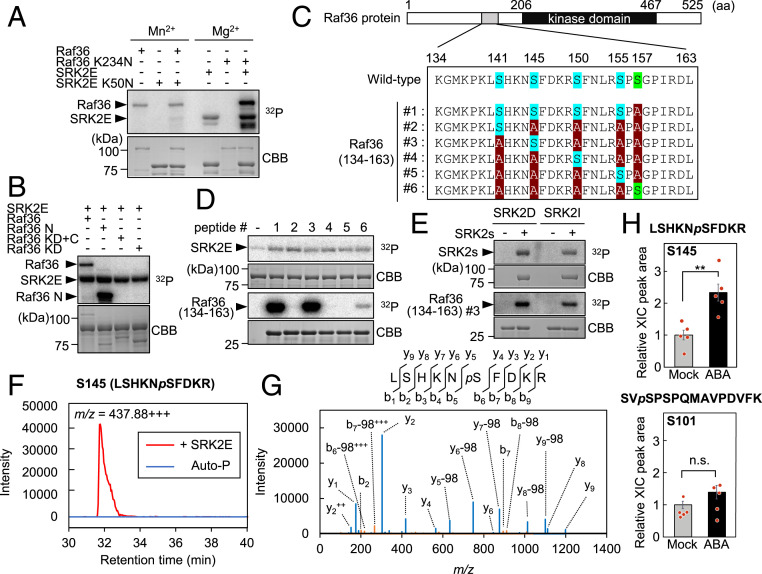
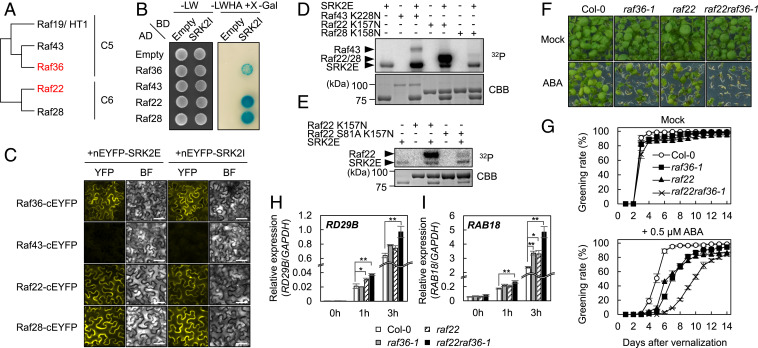
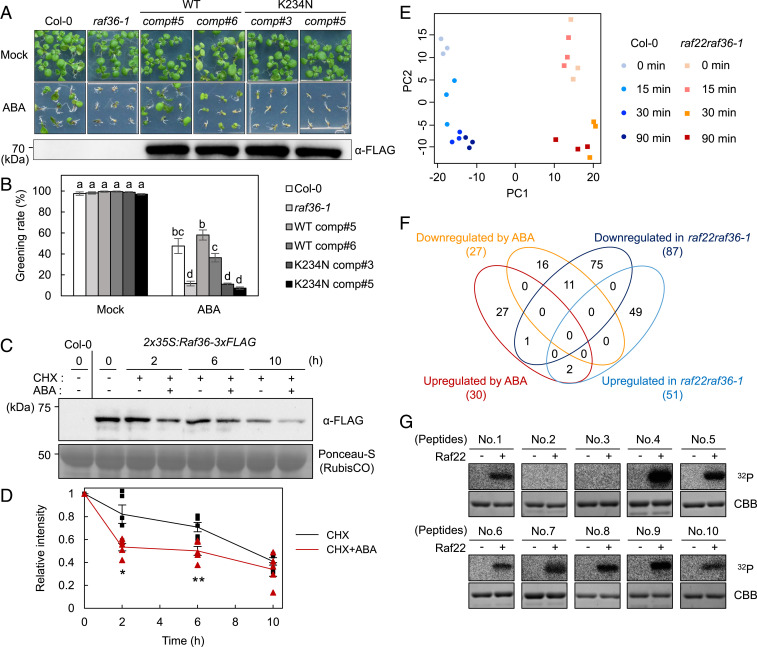
The phytohormone abscisic acid (ABA) plays a major role in abiotic stress responses in plants, and subclass III SNF1-related protein kinase 2 (SnRK2) kinases mediate ABA signaling. In this study, we identified Raf36, a group C Raf-like protein kinase in Arabidopsis, as a protein that interacts with multiple SnRK2s. A series of reverse genetic and biochemical analyses revealed that 1) Raf36 negatively regulates ABA responses during postgermination growth, 2) the N terminus of Raf36 is directly phosphorylated by SnRK2s, and 3) Raf36 degradation is enhanced in response to ABA. In addition, Raf22, another C-type Raf-like kinase, functions partially redundantly with Raf36 to regulate ABA responses. A comparative phosphoproteomic analysis of ABA-induced responses of wild-type and raf22raf36-1 plants identified proteins that are phosphorylated downstream of Raf36 and Raf22 in planta. Together, these results support a model in which Raf36/Raf22 function mainly under optimal conditions to suppress ABA responses, whereas in response to ABA, the SnRK2 module promotes Raf36 degradation as a means of alleviating Raf36-dependent inhibition and allowing for heightened ABA signaling to occur.
Keywords: Arabidopsis thaliana; Raf-like kinase; SnRK2; abscisic acid.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Shinozaki K., Yamaguchi-Shinozaki K., Seki M., Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 6, 410–417 (2003). - PubMed
-
- Cutler S. R., Rodriguez P. L., Finkelstein R. R., Abrams S. R., Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 61, 651–679 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
