

Insulin protects acinar cells during pancreatitis by preserving glycolytic ATP supply to calcium pumps
- PMID: 34282152
- PMCID: PMC8289871
- DOI: 10.1038/s41467-021-24506-w
Insulin protects acinar cells during pancreatitis by preserving glycolytic ATP supply to calcium pumps
Abstract
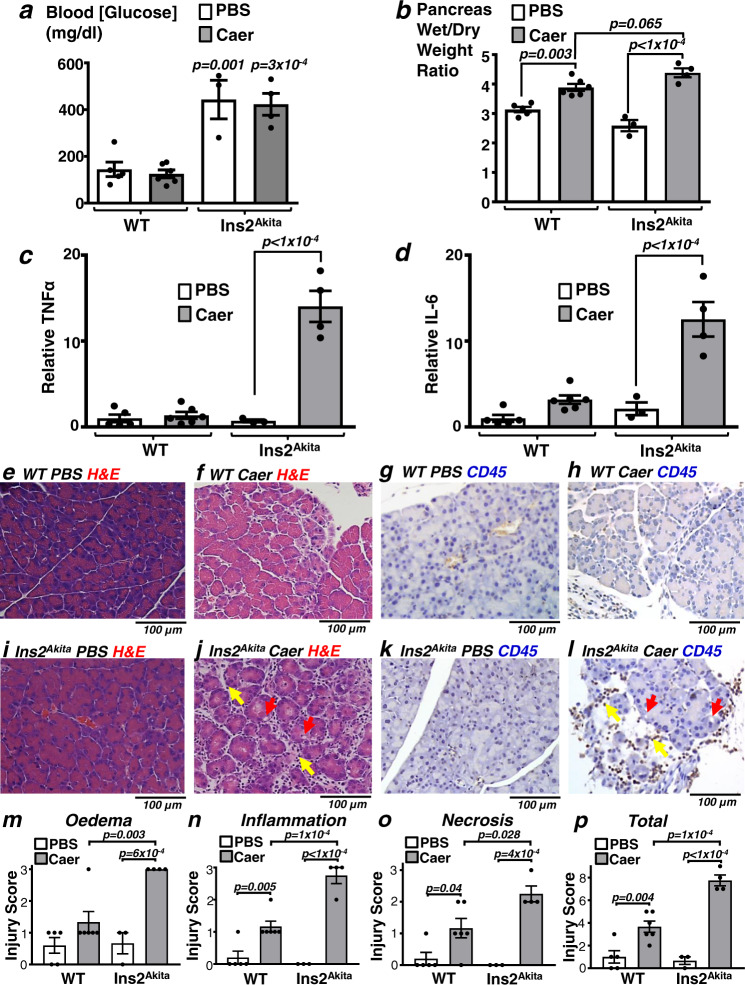
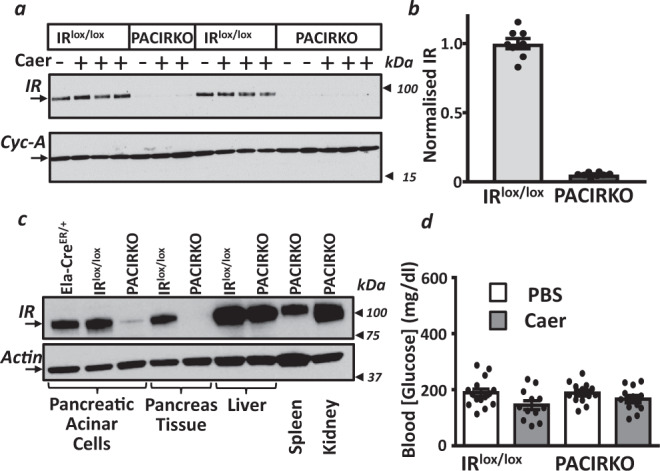
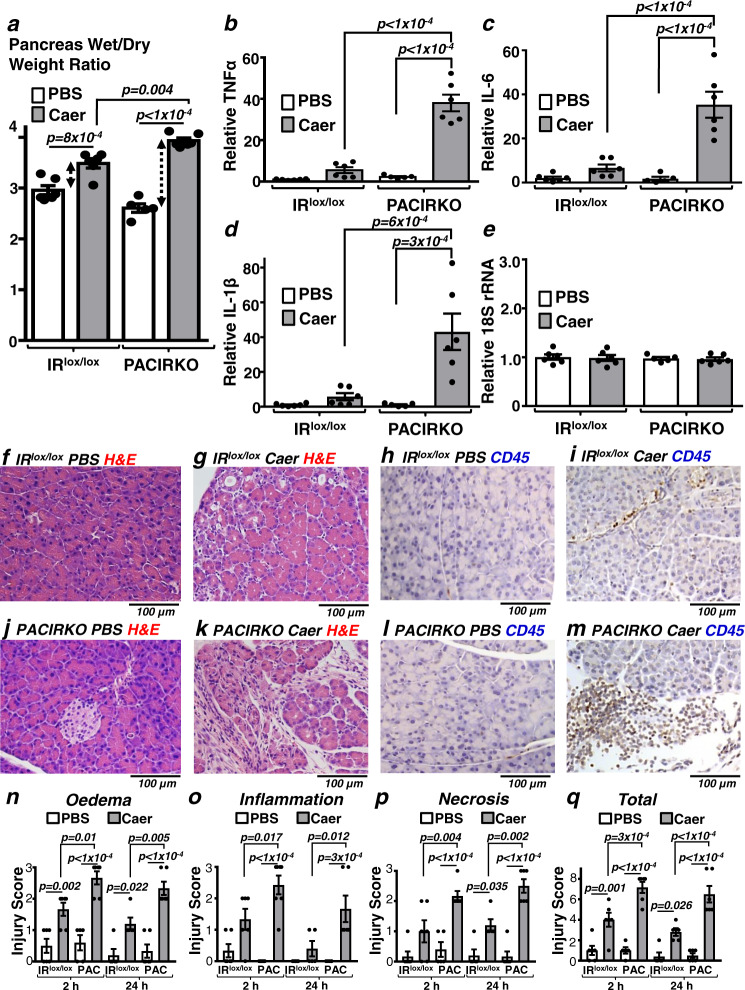
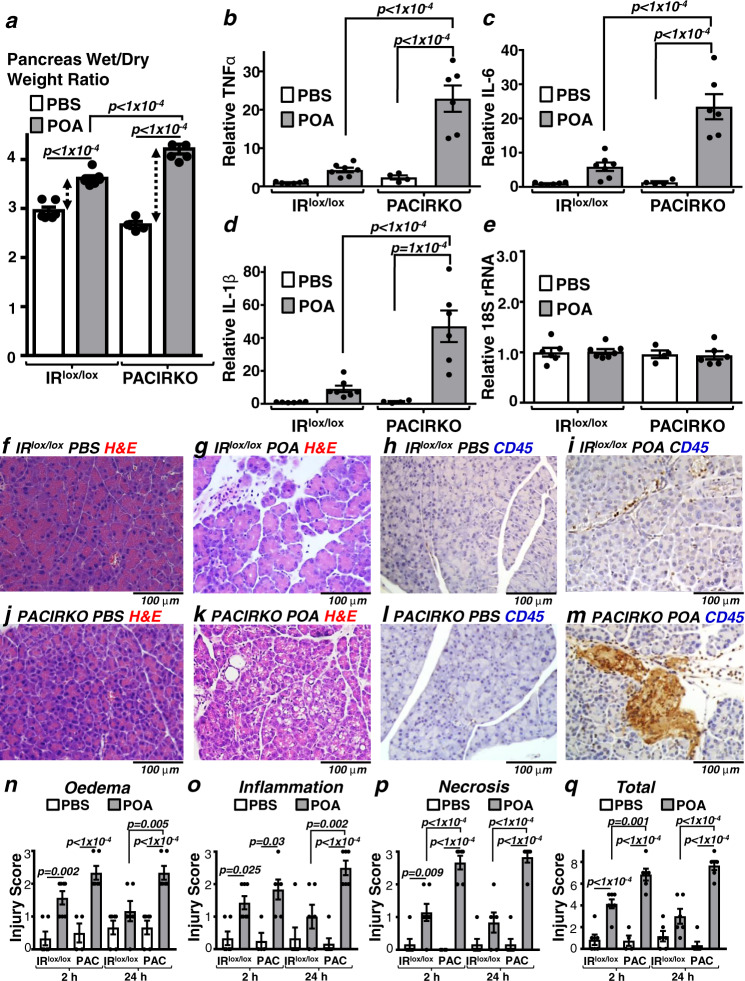
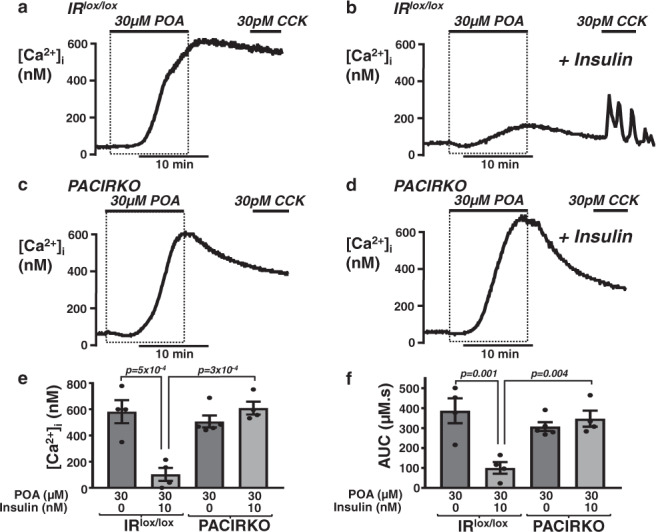
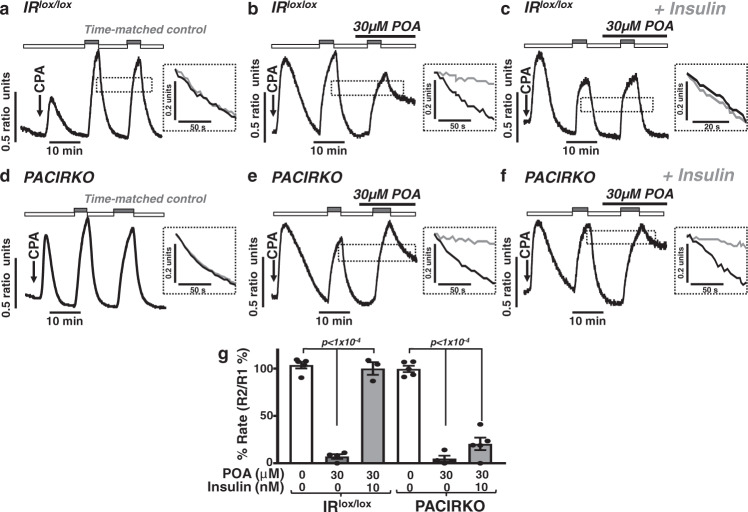
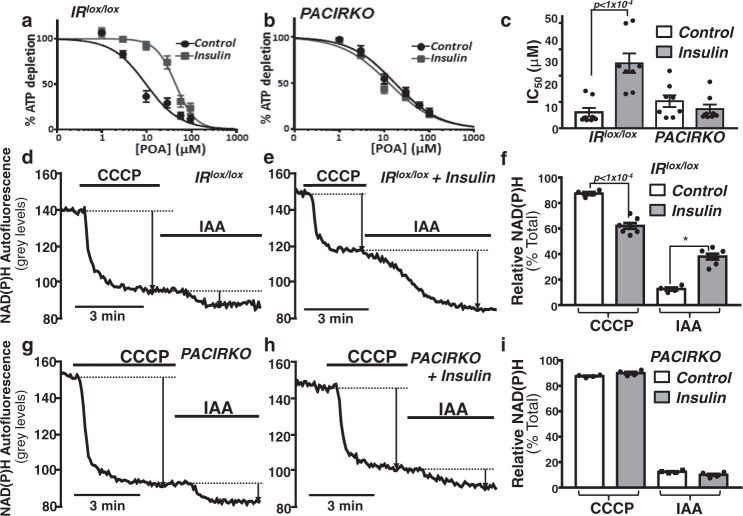
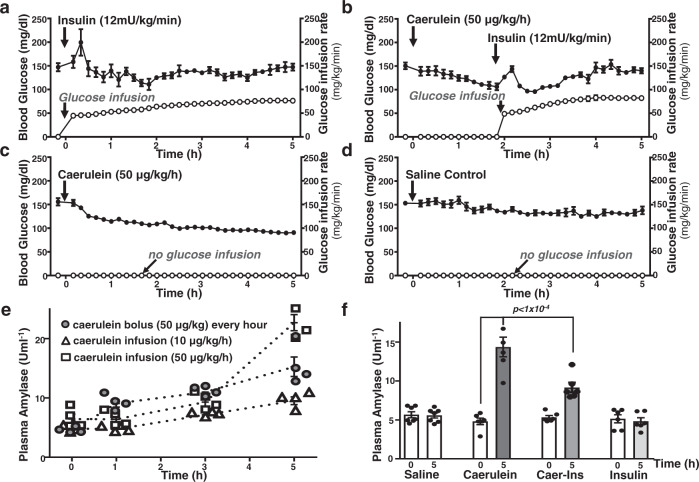
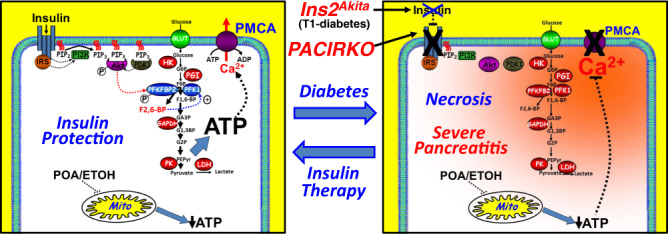
Acute pancreatitis (AP) is serious inflammatory disease of the pancreas. Accumulating evidence links diabetes with severity of AP, suggesting that endogenous insulin may be protective. We investigated this putative protective effect of insulin during cellular and in vivo models of AP in diabetic mice (Ins2Akita) and Pancreatic Acinar cell-specific Conditional Insulin Receptor Knock Out mice (PACIRKO). Caerulein and palmitoleic acid (POA)/ethanol-induced pancreatitis was more severe in both Ins2Akita and PACIRKO vs control mice, suggesting that endogenous insulin directly protects acinar cells in vivo. In isolated pancreatic acinar cells, insulin induced Akt-mediated phosphorylation of 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 2 (PFKFB2) which upregulated glycolysis thereby preventing POA-induced ATP depletion, inhibition of the ATP-dependent plasma membrane Ca2+ ATPase (PMCA) and cytotoxic Ca2+ overload. These data provide the first mechanistic link between diabetes and severity of AP and suggest that phosphorylation of PFKFB2 may represent a potential therapeutic strategy for treatment of AP.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
A protective role for insulin in acute pancreatitis.Nat Rev Endocrinol. 2021 Oct;17(10):577. doi: 10.1038/s41574-021-00547-5. Nat Rev Endocrinol. 2021. PMID: 34316042 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
