

Extracellular detection of neuronal coupling
- PMID: 34282275
- PMCID: PMC8289866
- DOI: 10.1038/s41598-021-94282-6
Extracellular detection of neuronal coupling
Abstract
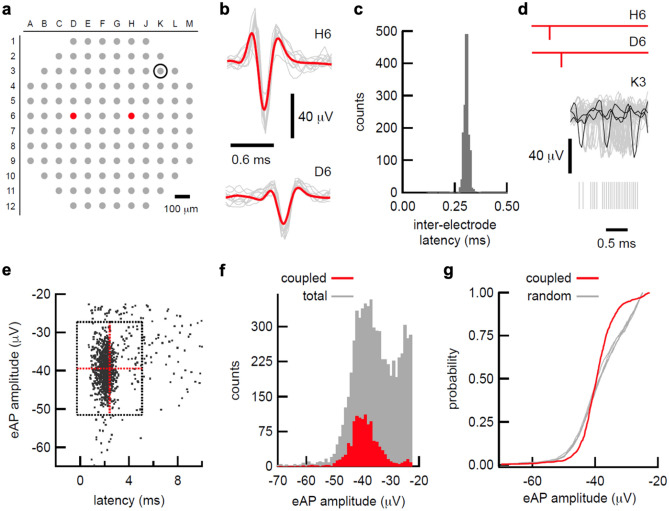
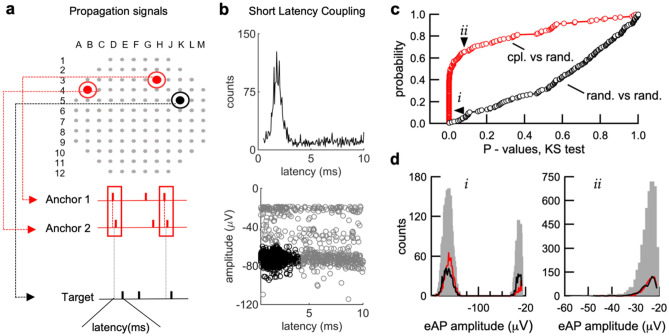
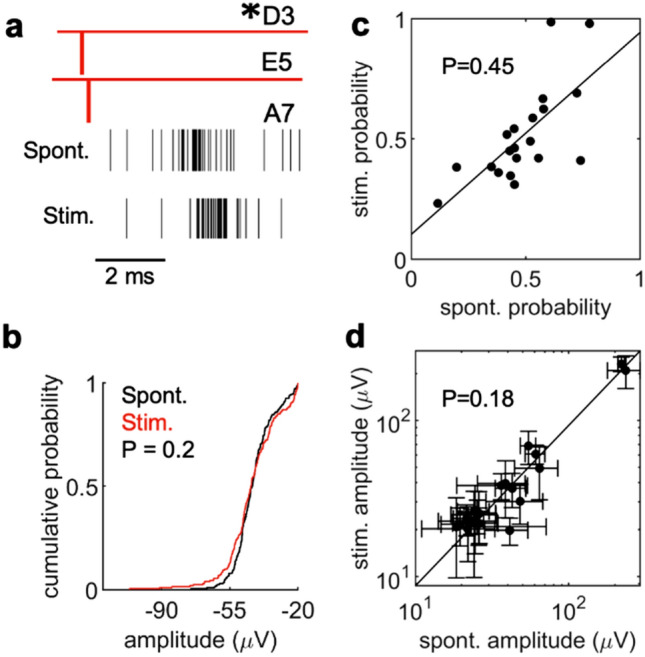
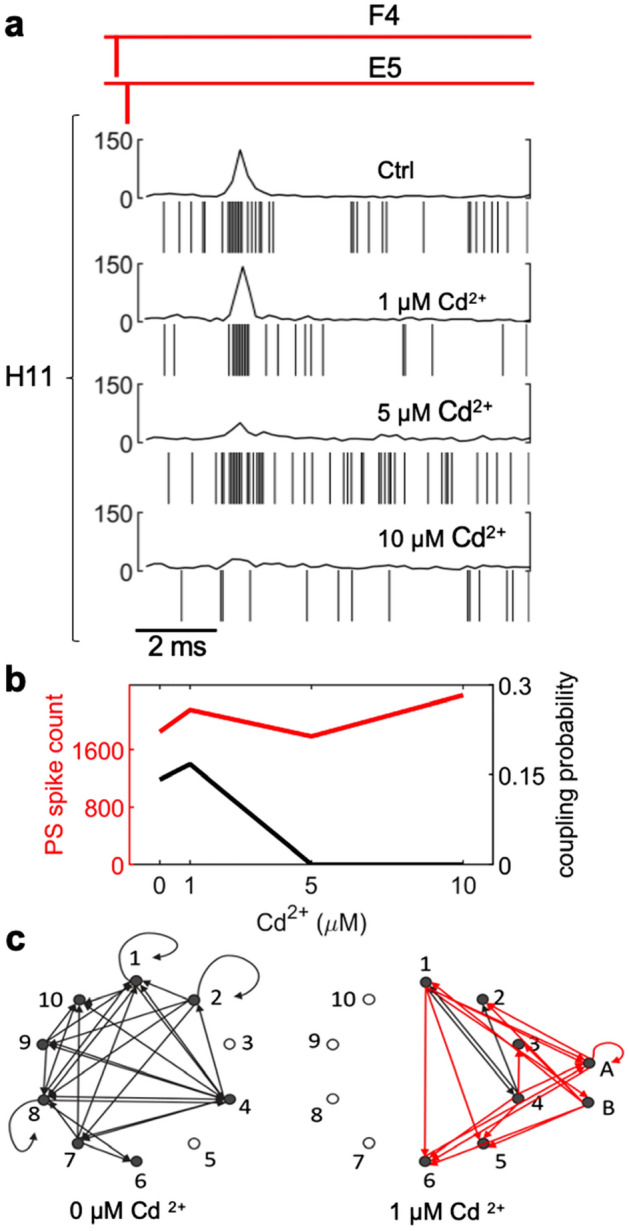
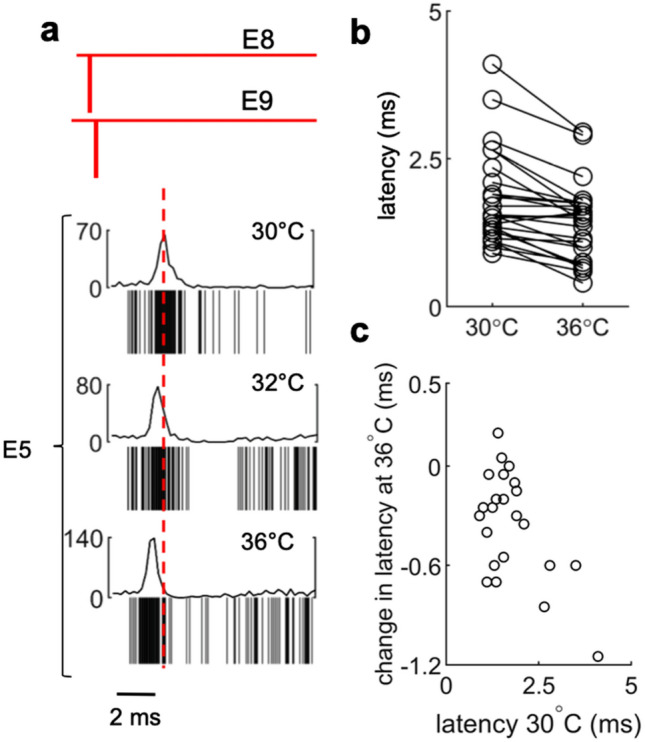
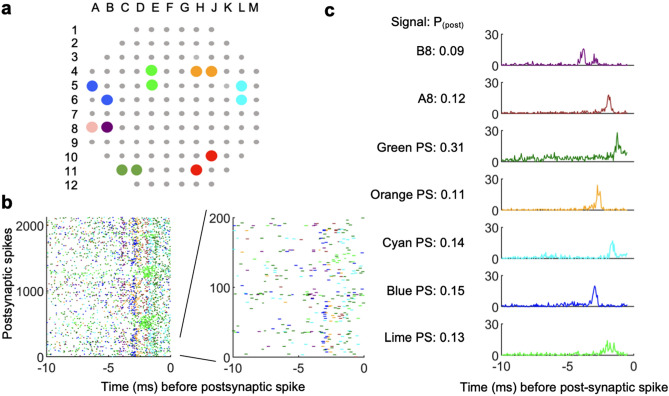
We developed a method to non-invasively detect synaptic relationships among neurons from in vitro networks. Our method uses microelectrode arrays on which neurons are cultured and from which propagation of extracellular action potentials (eAPs) in single axons are recorded at multiple electrodes. Detecting eAP propagation bypasses ambiguity introduced by spike sorting. Our methods identify short latency spiking relationships between neurons with properties expected of synaptically coupled neurons, namely they were recapitulated by direct stimulation and were sensitive to changing the number of active synaptic sites. Our methods enabled us to assemble a functional subset of neuronal connectivity in our cultures.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Action potential propagation recorded from single axonal arbors using multielectrode arrays.J Neurophysiol. 2018 Jul 1;120(1):306-320. doi: 10.1152/jn.00659.2017. Epub 2018 Apr 11. J Neurophysiol. 2018. PMID: 29641308
-
Extracellular recordings from patterned neuronal networks using planar microelectrode arrays.IEEE Trans Biomed Eng. 2004 Sep;51(9):1640-8. doi: 10.1109/TBME.2004.827252. IEEE Trans Biomed Eng. 2004. PMID: 15376512
-
A system for MEA-based multisite stimulation.IEEE Trans Biomed Eng. 2003 Feb;50(2):241-8. doi: 10.1109/TBME.2002.805470. IEEE Trans Biomed Eng. 2003. PMID: 12665038
-
Recent progress in multi-electrode spike sorting methods.J Physiol Paris. 2016 Nov;110(4 Pt A):327-335. doi: 10.1016/j.jphysparis.2017.02.005. Epub 2017 Mar 2. J Physiol Paris. 2016. PMID: 28263793 Free PMC article. Review.
-
Scaling Spike Detection and Sorting for Next-Generation Electrophysiology.Adv Neurobiol. 2019;22:171-184. doi: 10.1007/978-3-030-11135-9_7. Adv Neurobiol. 2019. PMID: 31073936 Review.
Cited by
-
From End to End: Gaining, Sorting, and Employing High-Density Neural Single Unit Recordings.Front Neuroinform. 2022 Jun 13;16:851024. doi: 10.3389/fninf.2022.851024. eCollection 2022. Front Neuroinform. 2022. PMID: 35769832 Free PMC article. Review.
