

NOP receptor antagonism attenuates reinstatement of alcohol-seeking through modulation of the mesolimbic circuitry in male and female alcohol-preferring rats
- PMID: 34285372
- PMCID: PMC8505627
- DOI: 10.1038/s41386-021-01096-1
NOP receptor antagonism attenuates reinstatement of alcohol-seeking through modulation of the mesolimbic circuitry in male and female alcohol-preferring rats
Abstract
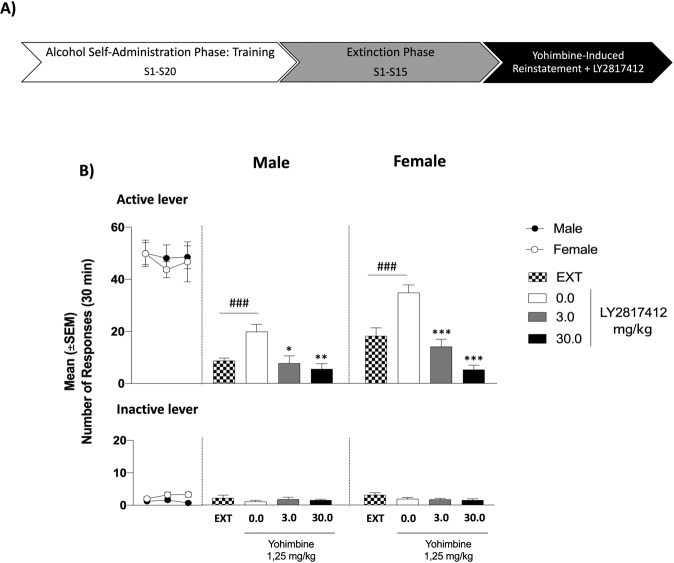
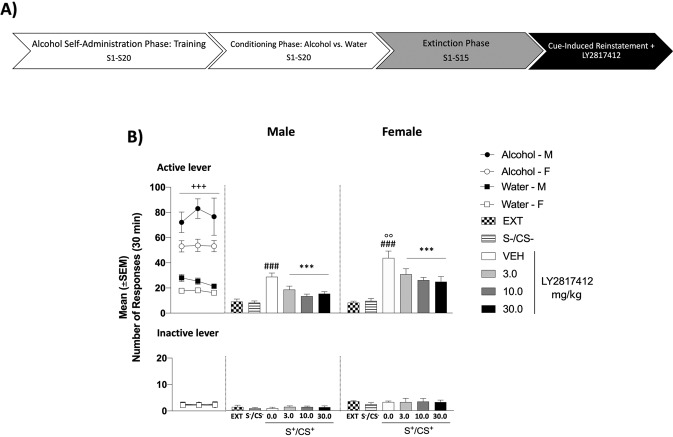
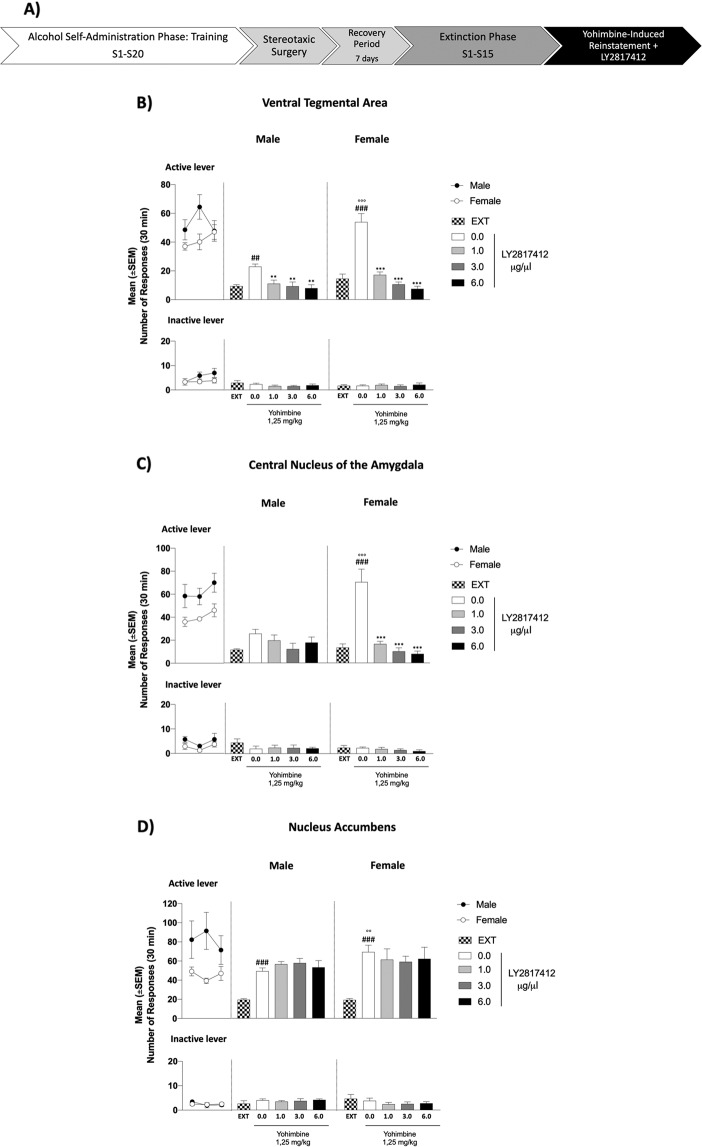
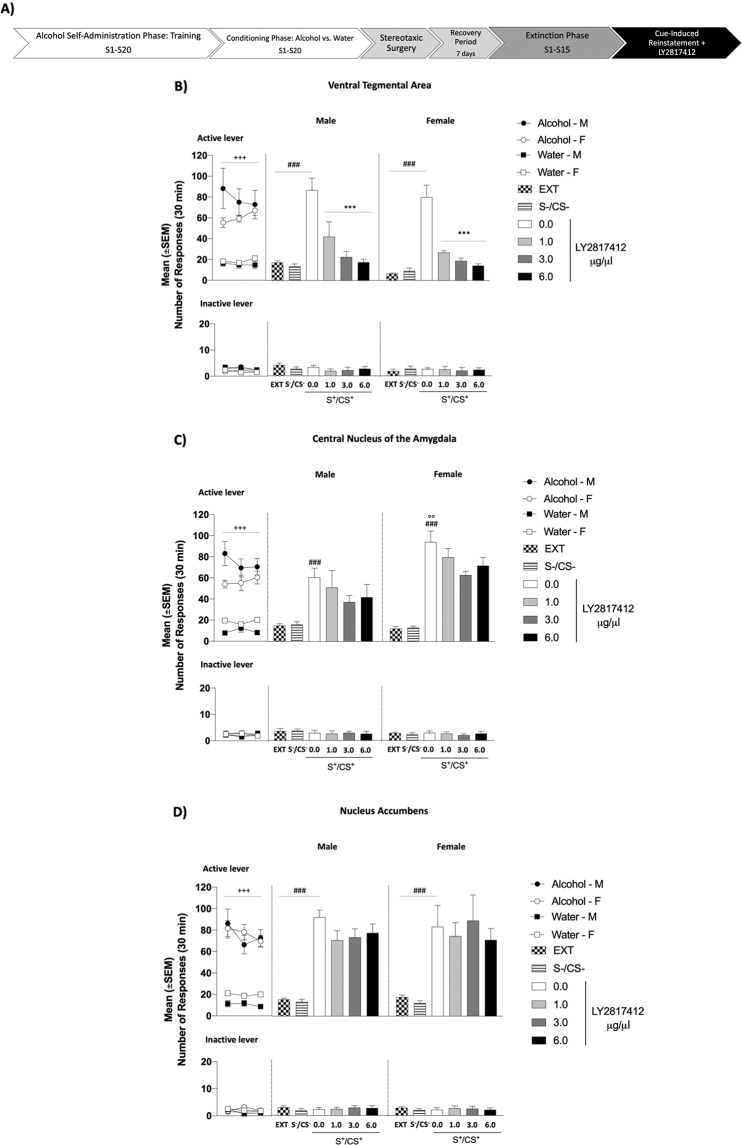
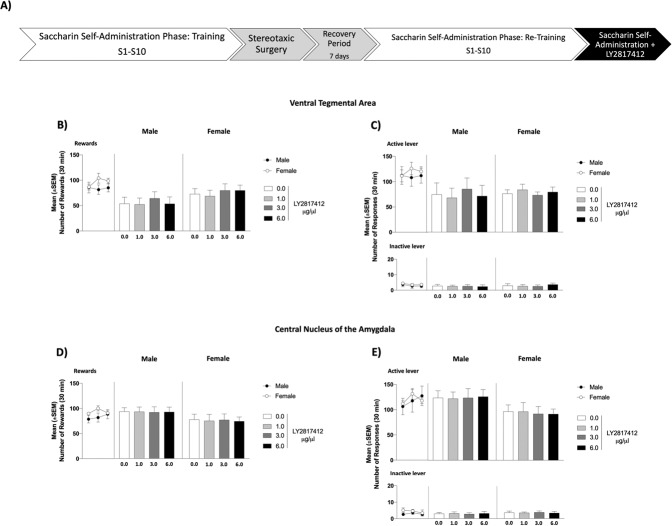
In patients suffering from alcohol use disorder (AUD), stress and environmental stimuli associated with alcohol availability are important triggers of relapse. Activation of the nociceptin opioid peptide (NOP) receptor by its endogenous ligand Nociceptin/Orphanin FQ (N/OFQ) attenuates alcohol drinking and relapse in rodents, suggesting that NOP agonists may be efficacious in treating AUD. Intriguingly, recent data demonstrated that also blockade of NOP receptor reduced alcohol drinking in rodents. To explore further the potential of NOP antagonism, we investigated its effects on the reinstatement of alcohol-seeking elicited by administration of the α2 antagonist yohimbine (1.25 mg/kg, i.p.) or by environmental conditioning factors in male and female genetically selected alcohol-preferring Marchigian Sardinian (msP) rats. The selective NOP receptor antagonist LY2817412 (0.0, 3.0, 10.0, and 30.0 mg/kg) was first tested following oral (p.o.) administration. We then investigated the effects of LY2817412 (1.0, 3.0, 6.0 μg/μl/rat) microinjected into three candidate mesolimbic brain regions: the ventral tegmental area (VTA), the central nucleus of the amygdala (CeA), and the nucleus accumbens (NAc). We found that relapse to alcohol seeking was generally stronger in female than in male rats and oral administration of LY2817412 reduced yohimbine- and cue-induced reinstatement in both sexes. Following site-specific microinjections, LY2817412 reduced yohimbine-induced reinstatement of alcohol-seeking when administered into the VTA and the CeA, but not in the NAc. Cue-induced reinstatement was suppressed only when LY2817412 was microinjected into the VTA. Infusions of LY2817412 into the VTA and the CeA did not alter saccharin self-administration. These results demonstrate that NOP receptor blockade prevents the reinstatement of alcohol-seeking through modulation of mesolimbic system circuitry, providing further evidence of the therapeutic potential of NOP receptor antagonism in AUD.
© 2021. The Author(s), under exclusive licence to American College of Neuropsychopharmacology.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Koob GF. Neurocircuitry of alcohol addiction: synthesis from animal models. Handb Clin Neurol. 2014;125:33–54. - PubMed
-
- Koob GF, Le Moal M. Plasticity of reward neurocircuitry and the ‘dark side’ of drug addiction. Nat Neurosci. 2005;8:1442–4. - PubMed
-
- Monti PM, Binkoff JA, Abrams DB, Zwick WR, Nirenberg TD, Liepman MR. Reactivity of alcoholics and nonalcoholics to drinking cues. J Abnorm Psychol. 1987;96:122–6. - PubMed
-
- Cooney NL, Litt MD, Morse PA, Bauer LO, Gaupp L. Alcohol cue reactivity, negative-mood reactivity, and relapse in treated alcoholic men. J Abnorm Psychol. 1997;106:243–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
