

Insertion of a synthetic switch into insulin provides metabolite-dependent regulation of hormone-receptor activation
- PMID: 34290145
- PMCID: PMC8325334
- DOI: 10.1073/pnas.2103518118
Insertion of a synthetic switch into insulin provides metabolite-dependent regulation of hormone-receptor activation
Abstract
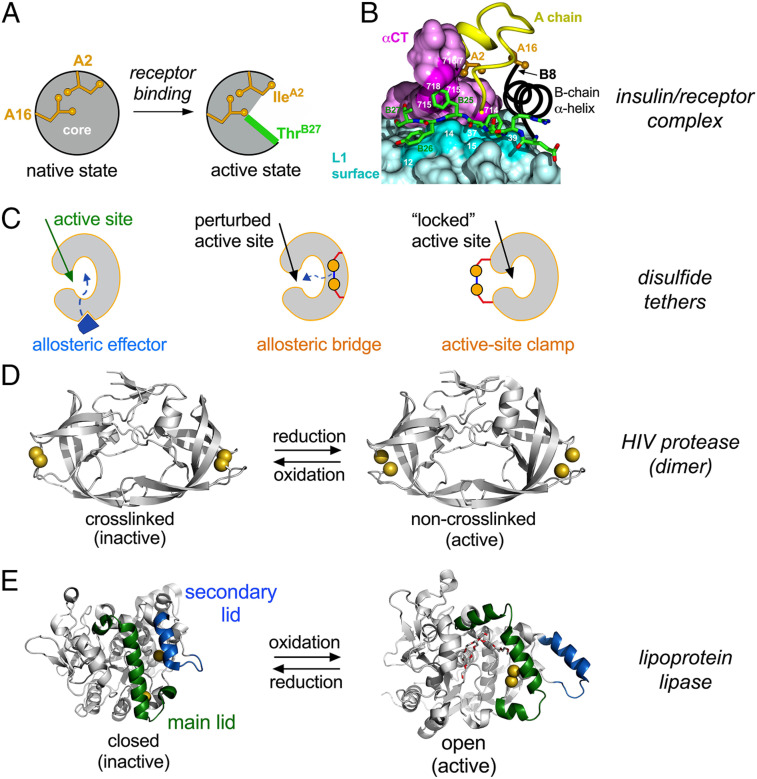
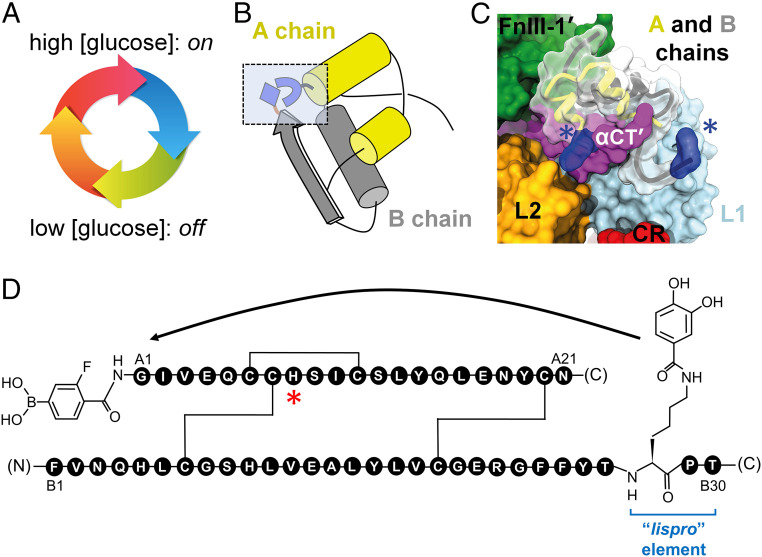
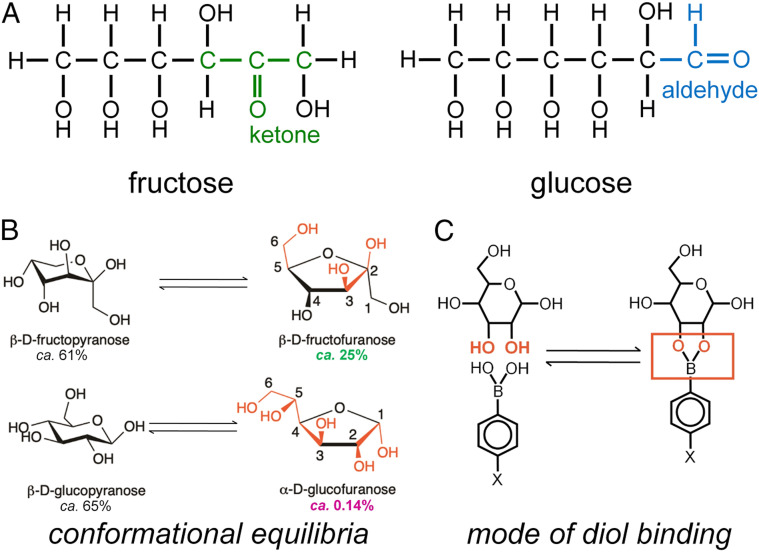
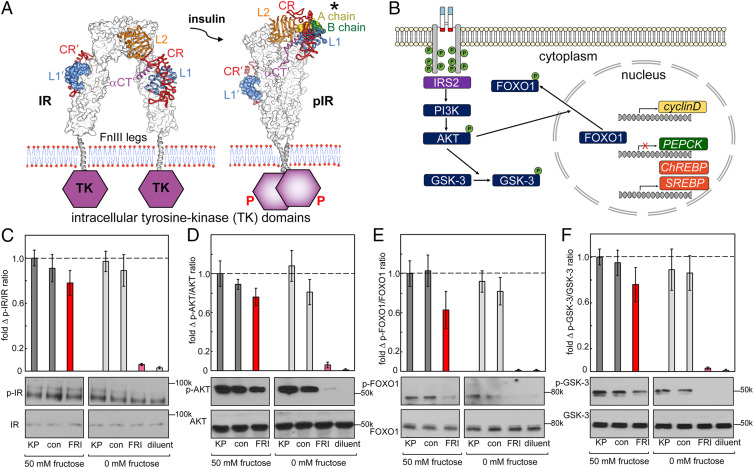
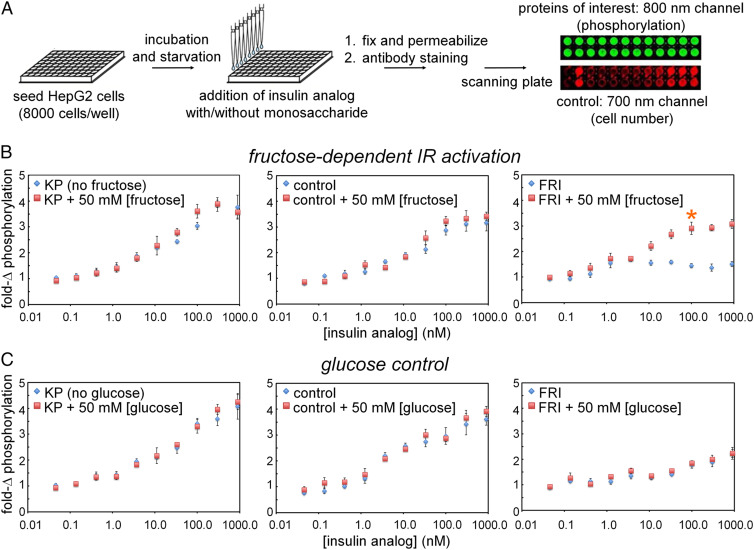
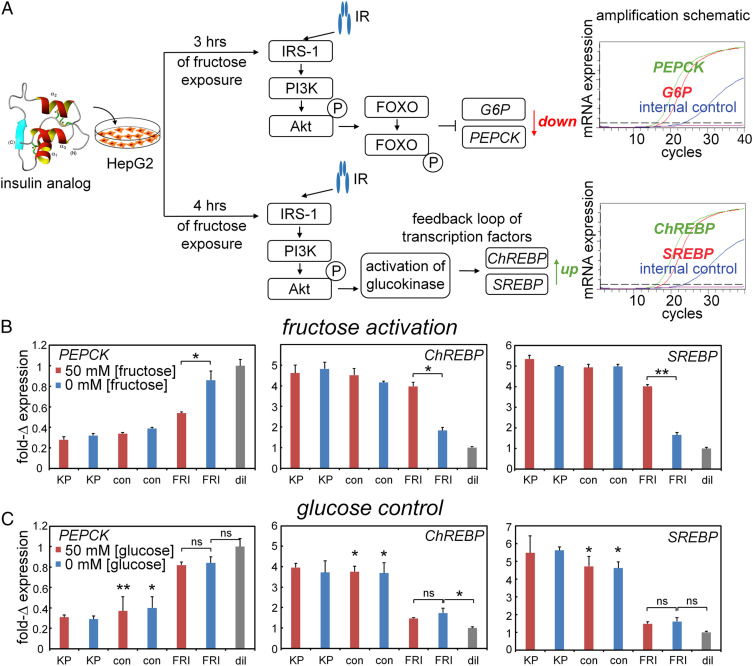
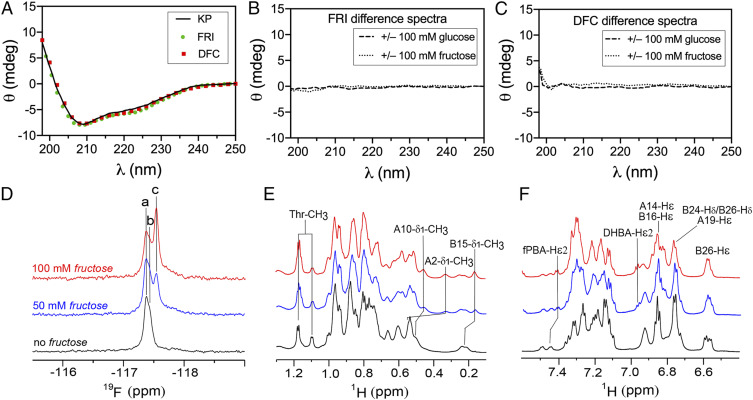
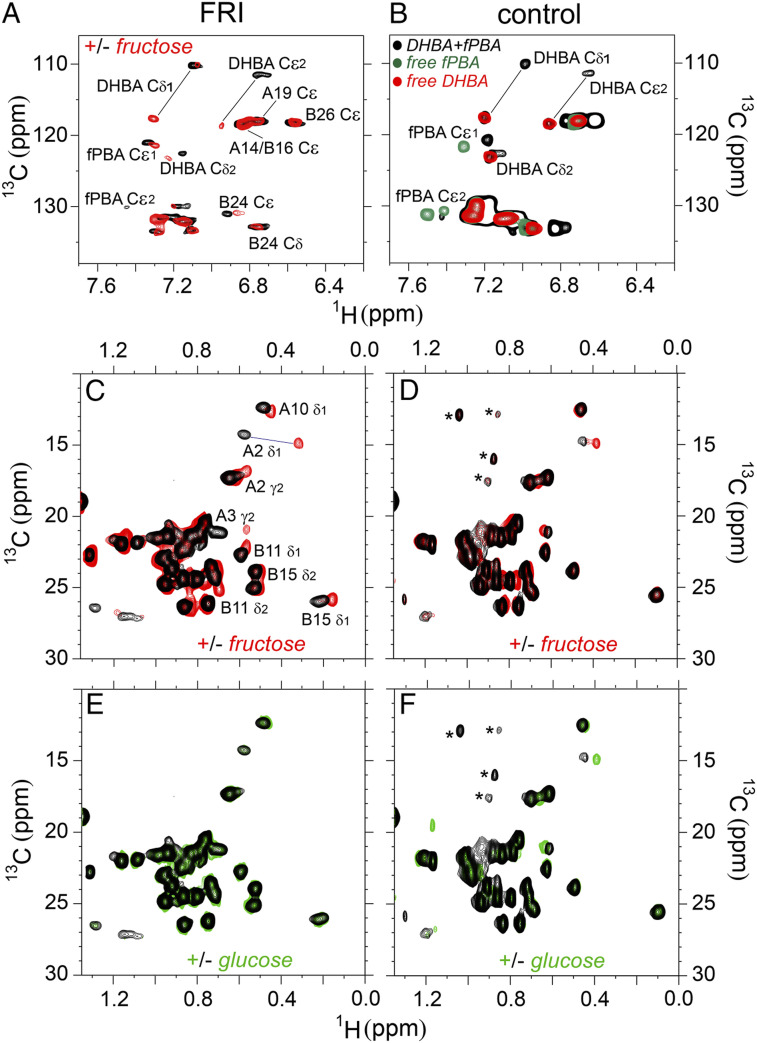
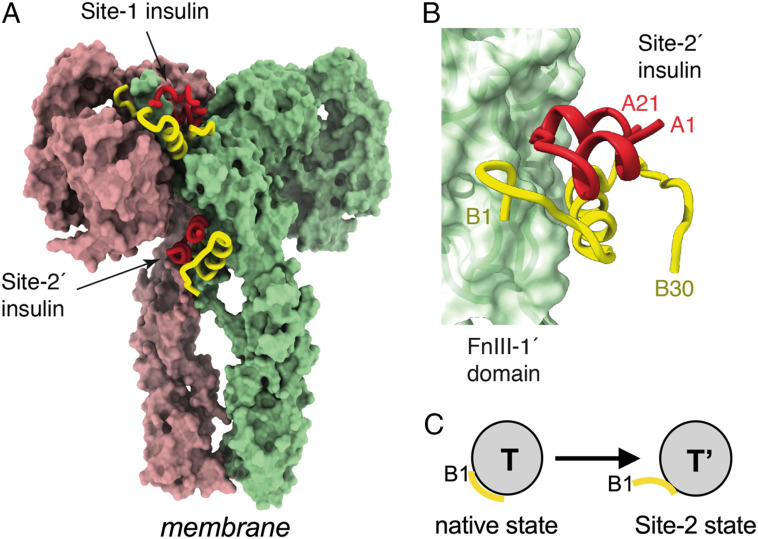
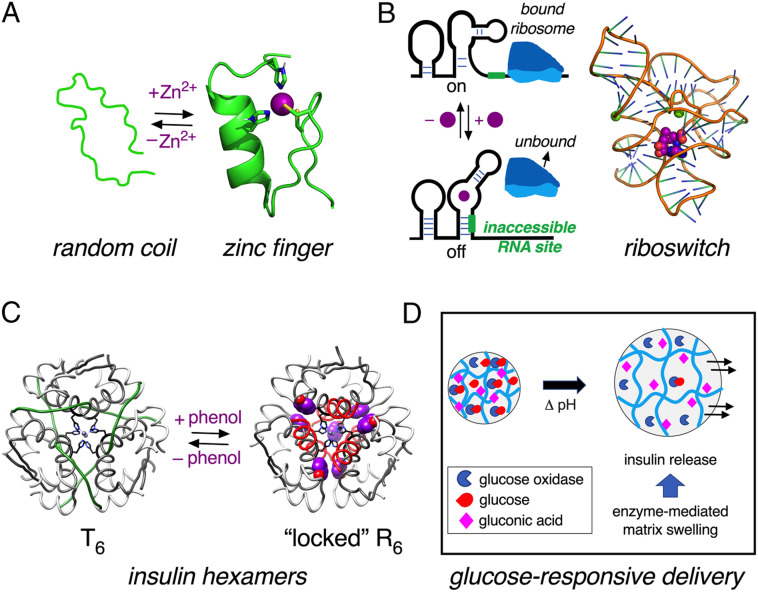
Insulin-signaling requires conformational change: whereas the free hormone and its receptor each adopt autoinhibited conformations, their binding leads to structural reorganization. To test the functional coupling between insulin's "hinge opening" and receptor activation, we inserted an artificial ligand-dependent switch into the insulin molecule. Ligand-binding disrupts an internal tether designed to stabilize the hormone's native closed and inactive conformation, thereby enabling productive receptor engagement. This scheme exploited a diol sensor (meta-fluoro-phenylboronic acid at GlyA1) and internal diol (3,4-dihydroxybenzoate at LysB28). The sensor recognizes monosaccharides (fructose > glucose). Studies of insulin-signaling in human hepatoma-derived cells (HepG2) demonstrated fructose-dependent receptor autophosphorylation leading to appropriate downstream signaling events, including a specific kinase cascade and metabolic gene regulation (gluconeogenesis and lipogenesis). Addition of glucose (an isomeric ligand with negligible sensor affinity) did not activate the hormone. Similarly, metabolite-regulated signaling was not observed in control studies of 1) an unmodified insulin analog or 2) an analog containing a diol sensor without internal tethering. Although secondary structure (as probed by circular dichroism) was unaffected by ligand-binding, heteronuclear NMR studies revealed subtle local and nonlocal monosaccharide-dependent changes in structure. Insertion of a synthetic switch into insulin has thus demonstrated coupling between hinge-opening and allosteric holoreceptor signaling. In addition to this foundational finding, our results provide proof of principle for design of a mechanism-based metabolite-responsive insulin. In particular, replacement of the present fructose sensor by an analogous glucose sensor may enable translational development of a "smart" insulin analog to mitigate hypoglycemic risk in diabetes therapy.
Keywords: diabetes mellitus; hormone–receptor recognition; insulin pharmacology; protein engineering; receptor tyrosine kinase.
Conflict of interest statement
Competing interest statement: M.A.W. has equity in Thermalin Inc. (Cleveland, OH) at which he serves as Chief Innovation Officer; he has also been a consultant to Merck Research Laboratories and DEKA Research and Development Corp. N.B.P. is a consultant to Thermalin Inc. M.C.L.'s laboratory has a Research Agreement with Eli Lilly and Company to conduct research not connected to this publication.
Figures
Comment in
References
-
- De Meyts P., Whittaker J., Structural biology of insulin and IGF1 receptors: Implications for drug design. Nat. Rev. Drug Discov. 1, 769–783 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
