

K+-dependent Müller cell-generated components of the electroretinogram
- PMID: 34294176
- PMCID: PMC10422678
- DOI: 10.1017/S0952523821000092
K+-dependent Müller cell-generated components of the electroretinogram
Abstract
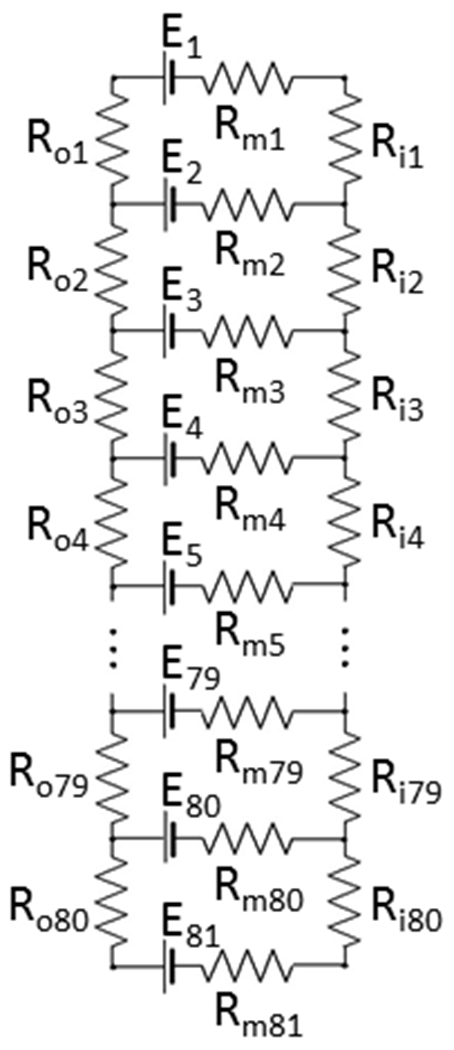
The electroretinogram (ERG) has been employed for years to collect information about retinal function and pathology. The usefulness of this noninvasive test depends on our understanding of the cell sources that generate the ERG. Important contributors to the ERG are glial Müller cells (MCs), which are capable of generating substantial transretinal potentials in response to light-induced changes in extracellular K+ concentration ([K+]o). For instance, the MCs generate the slow PIII (sPIII) component of the ERG as a reaction to a photoreceptor-induced [K+]o decrease in the subretinal space. Similarly, an increase of [K+]o related to activity of postreceptor retinal neurons also produces transretinal glial currents, which can potentially influence the amplitude and shape of the b-wave, one of the most frequently analyzed ERG components. Although it is well documented that the majority of the b-wave originates from On-bipolar cells, some contribution from MCs was suggested many years ago and has never been experimentally rejected. In this work, detailed information about light-evoked [K+]o changes in the isolated mouse retina was collected and then analyzed with a relatively simple linear electrical model of MCs. The results demonstrate that the cornea-positive potential generated by MCs is too small to contribute noticeably to the b-wave. The analysis also explains why MCs produce the large cornea-negative sPIII subcomponent of the ERG, but no substantial cornea-positive potential.
Keywords: K+-selective microelectrodes; Müller cells; computer simulation; electroretinogram; retina.
Figures








References
-
- Berger A, Cavallero S, Dominguez E, Barbe P, Simonutti M, Sahel J-A, Sennlaub F, Raoul W, Paques M & Bemelmans A-P (2014). Spectral-domain optical coherence tomography of the rodent eye: highlighting layers of the outer retina using signal averaging and comparison with histology. PloS one 9. - PMC - PubMed
-
- Bolnick DA, Walter AE & Sillman AJ (1979). Barium suppresses slow PIII in perfused bullfrog retina. Vision research 19, 1117–1119. - PubMed
-
- Brew H, Gray PT, Mobbs P & Attwell D (1986). Endfeet of retinal glial cells have higher densities of ion channels that mediate K+ buffering. Nature 324, 466. - PubMed
-
- Bringmann A, Pannicke T, Grosche J, Francke M, Wiedemann P, Skatchkov SN, Osborne NN & Reichenbach A (2006). Müller cells in the healthy and diseased retina. Progress in retinal and eye research 25, 397–424. - PubMed
-
- Brown KT (1968). The electroretinogram: its components and their origins. Vision research 8, 633–677. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
