

Mantle cell lymphoma polarizes tumor-associated macrophages into M2-like macrophages, which in turn promote tumorigenesis
- PMID: 34297045
- PMCID: PMC8341355
- DOI: 10.1182/bloodadvances.2020003871
Mantle cell lymphoma polarizes tumor-associated macrophages into M2-like macrophages, which in turn promote tumorigenesis
Abstract
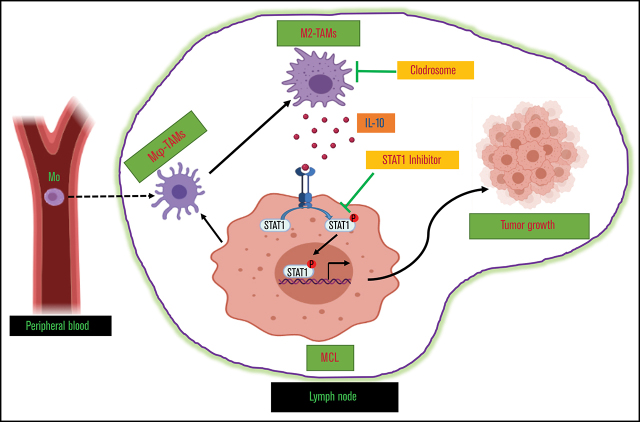
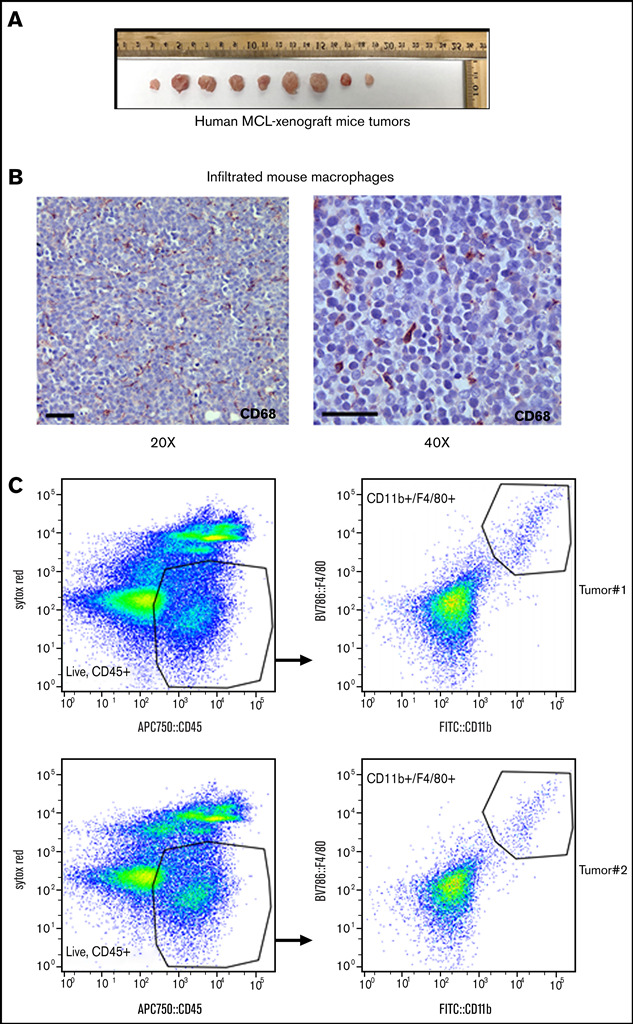
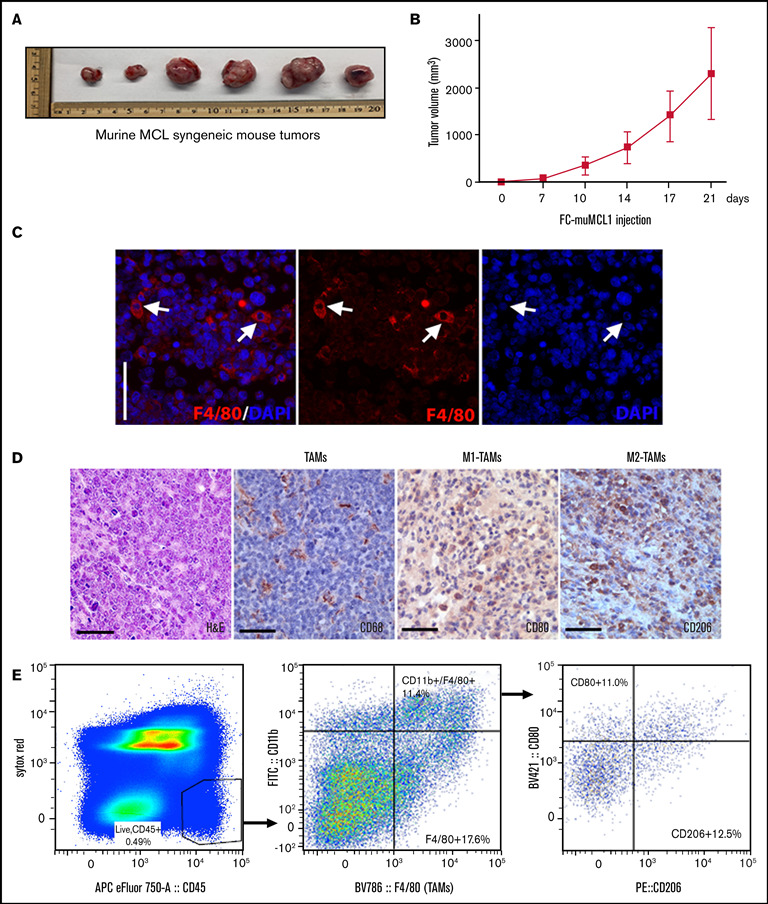
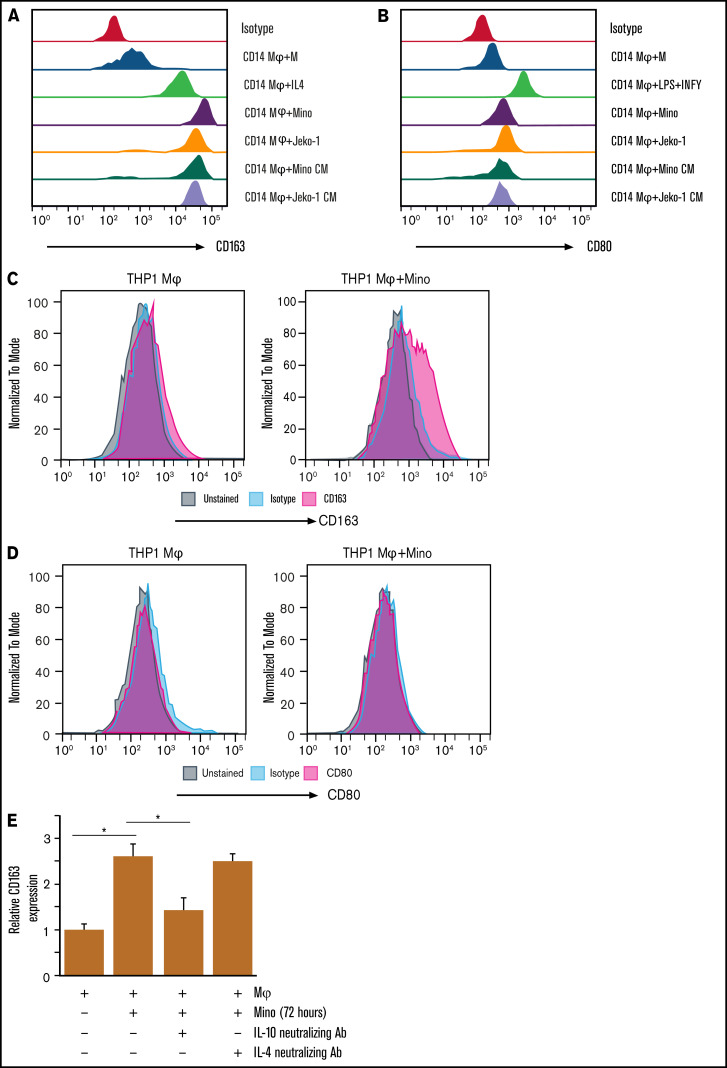
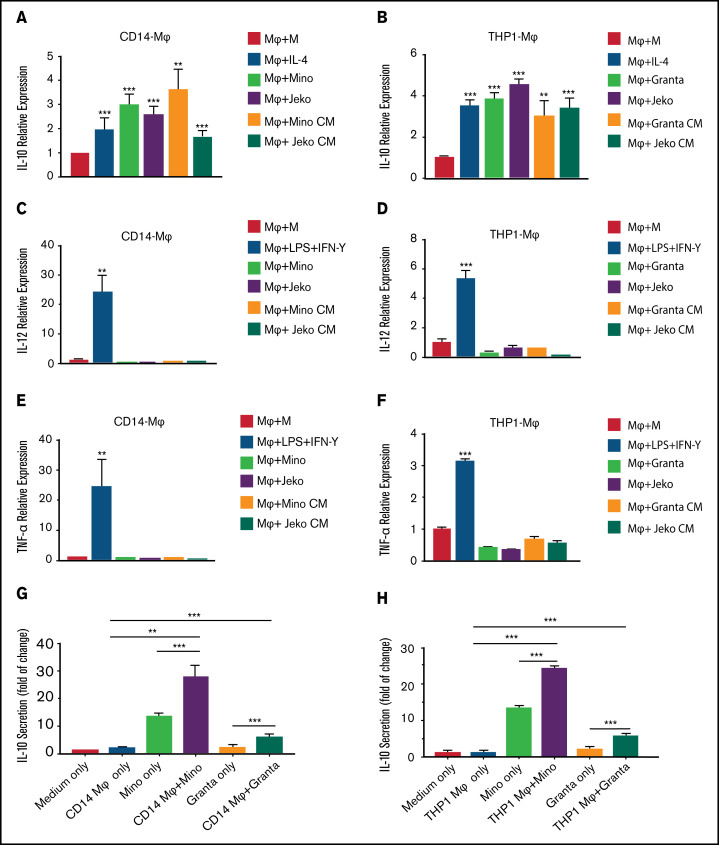
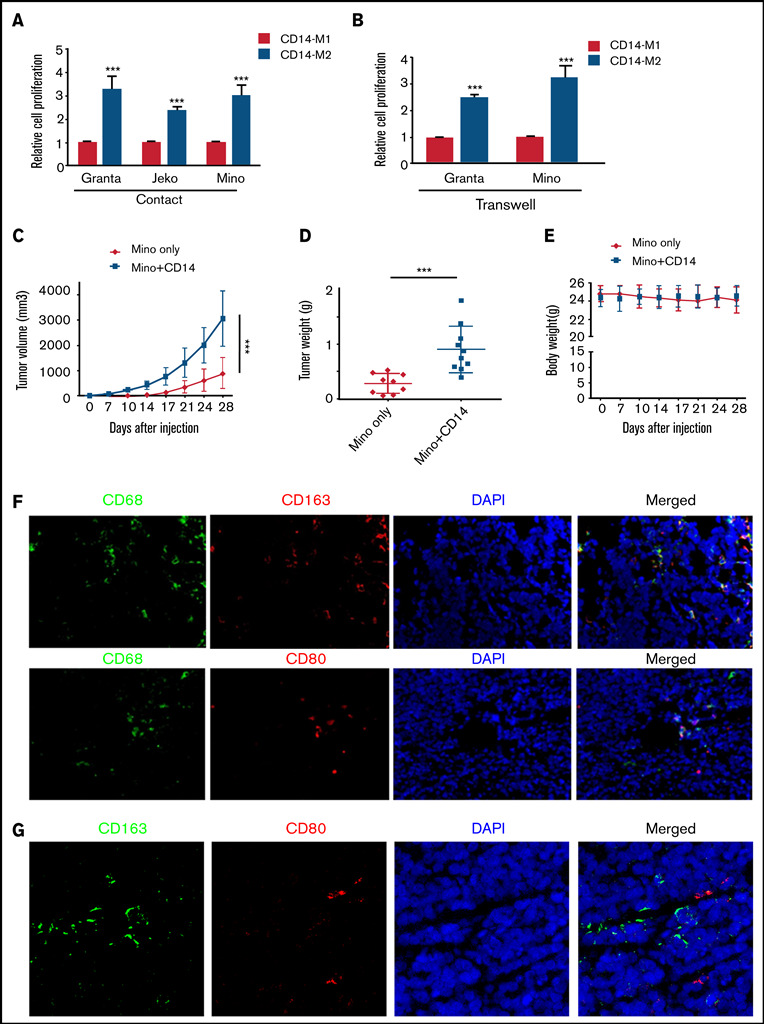
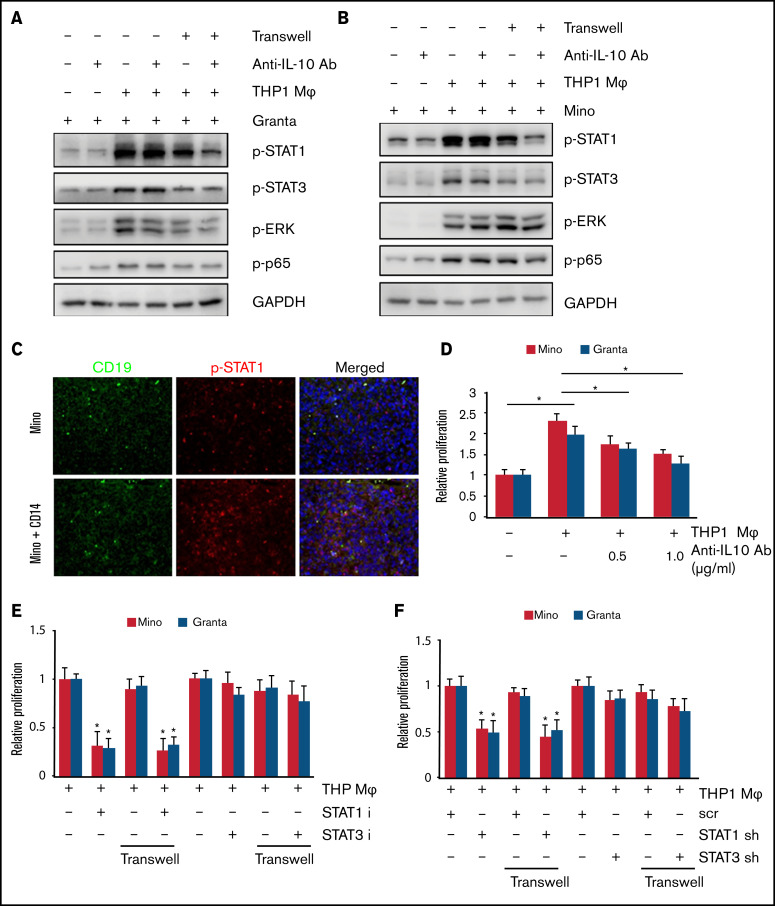
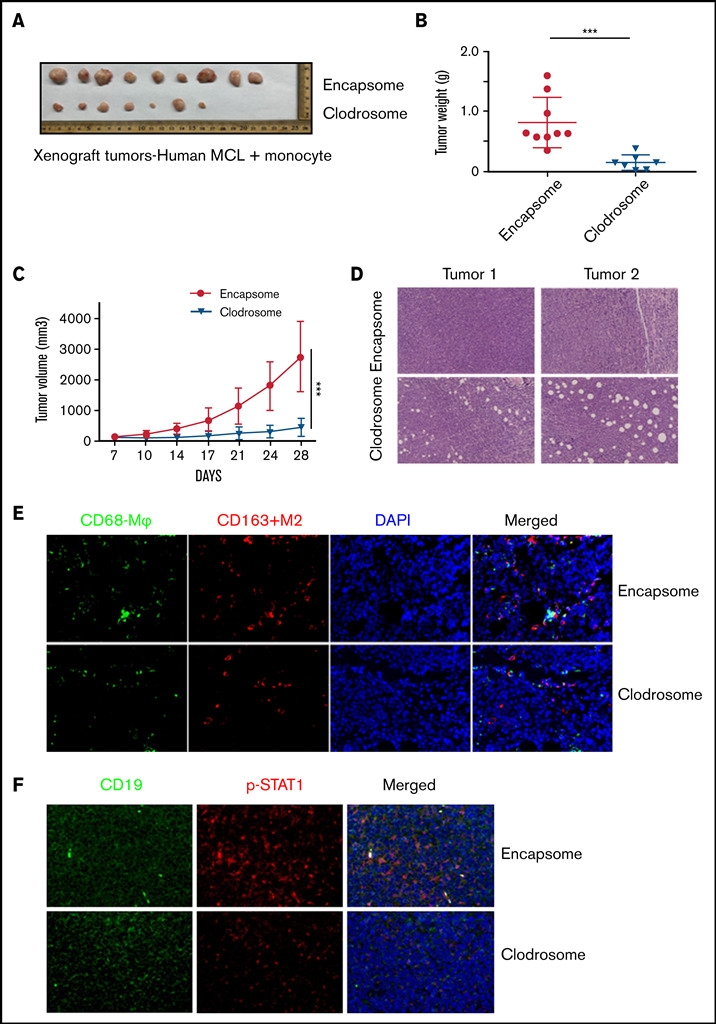
Tumor-associated macrophages (TAMs) are recognized as a hallmark of certain solid cancers and predictors of poor prognosis; however, the functional role of TAMs in lymphoid malignancies, including B-cell lymphoma, has not been well defined. We identified infiltration of F4/80+ TAMs in a syngeneic mouse model using the recently generated murine mantle cell lymphoma (MCL) cell line FC-muMCL1. Multicolor flow cytometric analysis of syngeneic lymphoma tumors showed distinct polarization of F4/80+ TAMs into CD206+ M2 and CD80+ M1 phenotypes. Using human MCL cell lines (Mino, Granta, and JVM2), we further showed that MCL cells polarized monocyte-derived macrophages toward an M2-like phenotype, as assessed by CD163+ expression and increased interleukin-10 (IL-10) level; however, levels of the M1 markers CD80 and IL-12 remained unaffected. To show that macrophages contribute to MCL tumorigenesis, we xenografted the human MCL cell line Mino along with CD14+ monocytes and compared tumor growth between these 2 groups. Results showed that xenografted Mino along with CD14+ monocytes significantly increased the tumor growth in vivo compared with MCL cells alone (P < .001), whereas treatment with liposomal clodronate (to deplete the macrophages) reversed the effect of CD14+ monocytes on growth of MCL xenografts (P < .001). Mechanistically, IL-10 secreted by MCL-polarized M2-like macrophages was found to be responsible for increasing MCL growth by activating STAT1 signaling, whereas IL-10 neutralizing antibody or STAT1 inhibition by fludarabine or STAT1 short hairpin RNA significantly abolished MCL growth (P < .01). Collectively, our data show the existence of a tumor microenvironmental network of macrophages and MCL tumor and suggest the importance of macrophages in interventional therapeutic strategies against MCL and other lymphoid malignancies.
© 2021 by The American Society of Hematology.
Figures
References
-
- Sandoval-Sus JD, Sotomayor EM, Shah BD.. Mantle cell lymphoma: contemporary diagnostic and treatment perspectives in the age of personalized medicine. Hematol Oncol Stem Cell Ther. 2017;10(3):99-115. - PubMed
-
- Rodgers TD, Reagan PM.. Targeting the B-cell receptor pathway: a review of current and future therapies for non-Hodgkin’s lymphoma. Expert Opin Emerg Drugs. 2018;23(2):111-122. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
