

Single cell functional genomics reveals plasticity of subcutaneous white adipose tissue (WAT) during early postnatal development
- PMID: 34298199
- PMCID: PMC8385178
- DOI: 10.1016/j.molmet.2021.101307
Single cell functional genomics reveals plasticity of subcutaneous white adipose tissue (WAT) during early postnatal development
Abstract
Objective: The current study addresses the cellular complexity and plasticity of subcutaneous (inguinal) white adipose tissue (iWAT) in mice during the critical periods of perinatal growth and establishment.
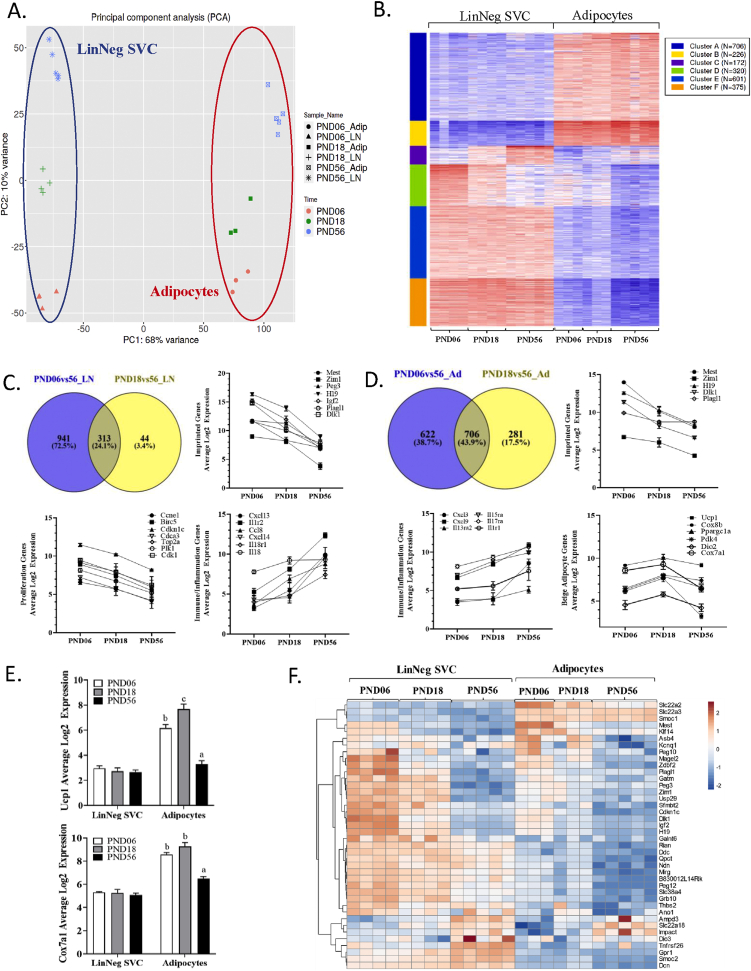
Methods: We performed a large-scale single cell transcriptomic (scRNA-seq) and epigenomic (snATAC-seq) characterization of cellular subtypes (adipose stromal cells (ASC) and adipocyte nuclei) during inguinal WAT (subcutaneous; iWAT) development in mice, capturing the early postnatal period (postnatal days (PND) 06 and 18) through adulthood (PND56).
Results: Perinatal and adult iWAT contain 3 major ASC subtypes that can be independently identified by RNA expression profiles and DNA transposase accessibility. Furthermore, the transcriptomes and enhancer landscapes of both ASC and adipocytes dynamically change during postnatal development. Perinatal ASC (PND06) are highly enriched for several imprinted genes (IGs; e.g., Mest, H19, Igf2) and extracellular matrix proteins whose expression then declines prior to weaning (PND18). By comparison, adult ASC (PND56) are more enriched for transcripts associated with immunoregulation, oxidative stress, and integrin signaling. Two clusters of mature adipocytes, identified through single nuclei RNA sequencing (snRNA-seq), were distinctive for proinflammatory/immune or metabolic gene expression patterns that became more transcriptionally diverse in adult animals. Single nuclei assay for transposase-accessible chromatin (snATAC-seq) revealed that differences in gene expression were associated with developmental changes in chromatin accessibility and predicted transcription factor motifs (e.g., Plagl1, Ar) in both stromal cells and adipocytes.
Conclusions: Our data provide new insights into transcriptional and epigenomic signaling networks important during iWAT establishment at a single cell resolution, with important implications for the field of metabolic programming.
Keywords: Adipocytes; Development; Imprinted genes; Single cell ATAC-Seq; Single cell RNA-Seq; White adipose tissue.
Copyright © 2021 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures





References
-
- Wang P., Mariman E., Renes J., Keijer J. The secretory function of adipocytes in the physiology of white adipose tissue. Journal of Cellular Physiology. 2008;216(1):3–13. - PubMed
-
- Bodis K., Roden M. Energy metabolism of white adipose tissue and insulin resistance in humans. European Journal of Clinical Investigation. 2018;48(11) - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
