

Antimelanogenesis Effects of Theasinensin A
- PMID: 34299073
- PMCID: PMC8305159
- DOI: 10.3390/ijms22147453
Antimelanogenesis Effects of Theasinensin A
Abstract
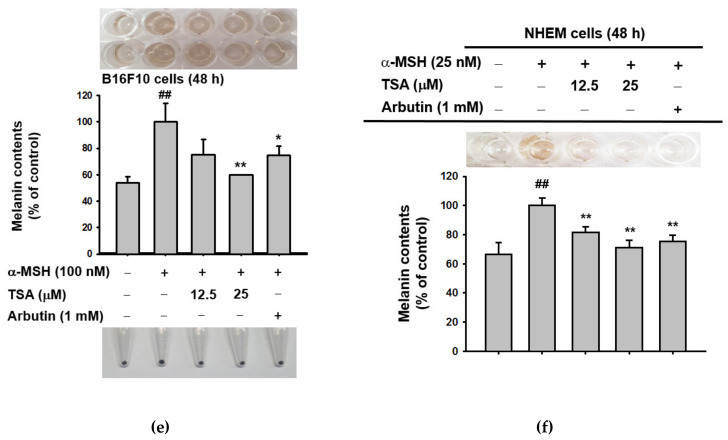
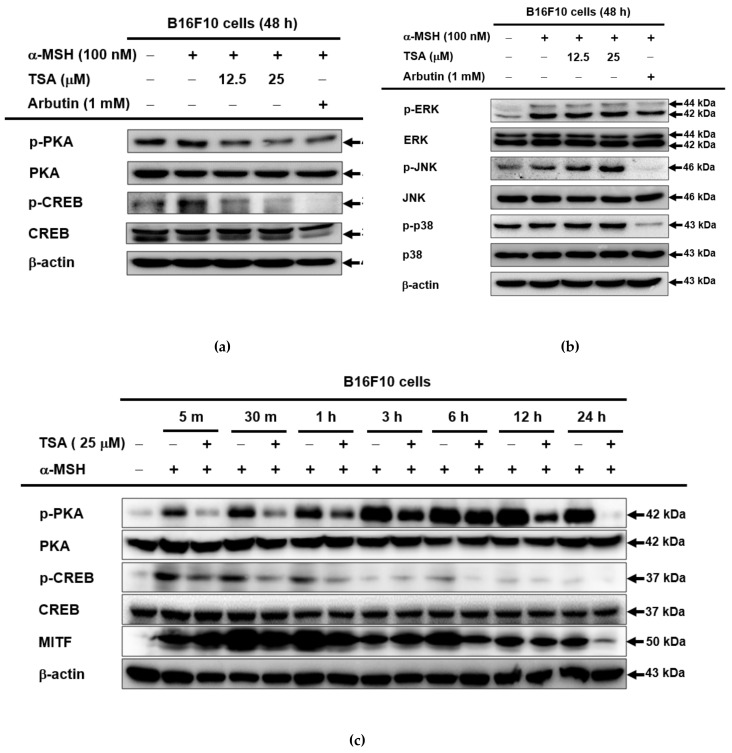
Theasinensin A (TSA) is a major group of catechin dimers mainly found in oolong tea and black tea. This compound is also manufactured with epigallocatechin gallate (EGCG) as a substrate and is refined after the enzyme reaction. In previous studies, TSA has been reported to be effective against inflammation. However, the effect of these substances on skin melanin formation remains unknown. In this study, we unraveled the role of TSA in melanogenesis using mouse melanoma B16F10 cells and normal human epidermal melanocytes (NHEMs) through reverse transcription polymerase chain reaction (RT-PCR), Western blotting analysis, luciferase reporter assay, and enzyme-linked immunosorbent assay analysis. TSA inhibited melanin formation and secretion in α-melanocyte stimulating hormone (α-MSH)-induced B16F10 cells and NHEMs. TSA down-regulated the mRNA expression of tyrosinase (Tyr), tyrosinase-related protein 1 (Tyrp1), and Tyrp2, which are all related to melanin formation in these cells. TSA was able to suppress the activities of certain proteins in the melanocortin 1 receptor (MC1R) signaling pathway associated with melanin synthesis in B16F10 cells: cyclic adenosine monophosphate (cAMP) response element-binding protein (CREB), protein kinase A (PKA), tyrosinase, and microphthalmia-associated transcription factor (MITF). We also confirmed α-MSH-mediated CREB activities through a luciferase reporter assay, and that the quantities of cAMP were reduced by TSA in the enzyme linked immunosorbent assay (ELISA) results. Based on these findings, TSA should be considered an effective inhibitor of hyperpigmentation.
Keywords: CREB; MC1R; cAMP; melanogenesis; theasinensin A.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
